|
Haubenformen bei Tauben
Als Grundformen der Haube werden meist die Spitzkappe und die Rund-
oder Muschelhaube unterschieden, die sich selbst in Details noch
voneinander unterscheiden. Das betrifft die Federfülle, bei der
Muschelhaube den seitlichen Abschluss durch eine Rosette oder nicht,
die Höhe des Ansatzes im Nacken u.a.

Grundformen von Federhauben: Spitzkappe, Rund- oder Muschelhaube
ohne Rosetten und Muschelhaube mit -seitlichen Rosetten, Fotos Layne
Gardner, Quelle: Pigeon Genetics.
Von zwölf auf Tafel 8 abgebildeten Tauben von
Aldrovandi in der
Frankfurter Ausgabe 1610 hatten fünf eine Spitzkappe. Eine weitere,
die Taube ‚mit dem Kamm‘ als vermuteter Vorläufer der Perückentaube,
hatte eine hochstehende Nackenmähne. Die anderen waren glattköpfig.
Tauben mit einer Rundhaube wurden in dem Werk nicht gezeigt.
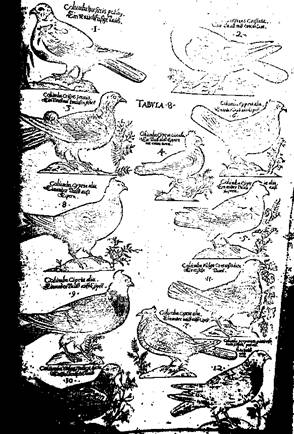
Taubenabbildungen bei Aldrovandi auf Bildtafel 8 der Frankfurter
Ausgabe von 1610
Auf den Bildtafeln des in Farsi abgefassten ‚Book of Pigeon‘ von
Musavi (um 1770) über
die Tauben aus dem Mogulreich vom 16. – 19. Jahrhundert auf dem
indischen Subkontinent gibt es Tauben mit Spitzkappe, aber keine mit
Rundhaube.
Meilensteine in der genetischen Analyse
Fast genau 300 Jahre nach der Darstellung von
Aldrovandi gab es 1911
eine erste wissenschaftliche genetische Analyse durch den späteren
Nobelpreisträger T.H. Morgan.
Morgan brachte früh die Frage von Federstrukturen wie Haube
und Schwanzfederzahlen in den Kontext der Mendelschen
Vererbungsgesetze. Er hatte u.a. eine glattköpfige Pfautaube und
eine rundhaubige Schwalbentaube verpaart und daraus 7 Jungtiere mit
Haube gezogen. Aus einem solchen Paar gab es in der F2
sowohl glattköpfige als auch behaubte Jungtiere. Nach den
Ergebnissen kam er zum Schluss eines dominanten Genes.

Erbversuche bei Morgan 1911. Quelle: Sell, Pigeon Genetics 2012,
Abb. 279.
Das wurde bald widerlegt. Die glattköpfige Pfautaube von
Morgan war
möglicherweise mischerbig und die Stichprobe wahrscheinlich zu
gering. Eine optisch fehlende Kappe kann auch durch eine schwache
Ausprägung erklärt werden, schließlich auch durch bei vielen
Merkmalen kaum erforschte epistatische Effekte mit ‚Enablern‘,
‚Verstärkern‘ oder blockierenden Faktoren.
Christie und Wriedt
dokumentierten 1923 durch umfangreiche Paarungen die rezessive
Vererbung der Rundhaube. Von 80 Jungtieren der F1 von
rundhaubigen Schildmövchen (Petenten) mit Dänischen glattköpfigen
Tümmlern waren alle glattköpfig. Von den 64 Jungtieren der F2
waren 44 glattköpfig, 17 rundhaubig und 3 spitzkappig. Auch
unter den 66 Rückpaarungen an Petenten gab es mit 4 Spitzkappigen
eine bei den Ausgangstieren nicht vorhandene Kappenform. Damit
rückten Unterschiede in der Federstruktur in den Fokus.
Morgan
hatte sich für die Unterschiede von Spitzkappe und Muschelhaube
nicht interessiert, obwohl er mit beiden Formen experimentiert
hatte. Soederberg
stellte 1927 für spitzkappige Gimpeltauben bei Paarungen mit
Glattköpfigen – wie Christie
und Wriedt für die Rundhaube der Petenten – die rezessive
Vererbung fest. Er ging noch einen Schritt weiter und verpaarte eine
rundhaubige Schwalbentaube mit einem Spitzkappigen und erhielt
daraus 6 rundhaubige Jungtiere, was er als Dominanz der Rundhaube
interpretierte. Christie und
Wriedt hatten dagegen als Ursache der Unterschiede
Modifikatoren vermutet, sind zu der angekündigten weiteren Analyse
aber nicht mehr gekommen.
Noch einmal etwa einhundert Jahre später wurde im Genom der
Haustaube ein Gen zur Ausbildung der Haube durch eine Forschergruppe
der University Utah identifiziert (M.D.
Shapiro u.a. 2013). Mit den Untersuchungen der
molekulargenetischen Grundlagen der Haustaube wurde für
Federstrukturen das Fenster zu einem anderen Untersuchungsdesign als
der optischen Auswertung von Phänotypen und Kreuzungsergebnissen
beschritten. Überraschend für Taubenzüchter an den Ergebnissen ist
nicht so sehr, dass sie einen Bereich im Genom identifiziert haben,
den alle getesteten Tiere mit einem befiederten Kopf gemeinsam
haben, sondern dass sie keinen Anzeichen für Allele des
identifizierten Genes und auch keine weiteren Bereiche finden
konnten, die Unterschiede bei Tieren mit Spitzkappe und Tieren mit
Muschelhaube aufwiesen und zur Erklärung beitragen könnten. In den
Zuchten vererben sowohl Spitzkappen und Muschelhauben jeweils rein
und zuverlässig und weisen in der Federstruktur deutliche
Unterschiede zueinander auf, so dass es verwunderlich erscheint,
wenn sich das in den DNS nicht widerspiegeln sollte.
Modifikatoren oder multiple Allele?
Spitzkappige nach der Verpaarung von Rundhaubigen mit Glattköpfigen
Eigene Erfahrungen mit der Verpaarung reinerbiger rundhaubiger
Tauben mit reinerbigen glattköpfigen Tauben zeigten, dass
spitzkappige Nachkommen in der F2 und in der R1
(erste Rückpaarung) nicht extrem selten sind.

Wenn der verantwortliche Modifikator im Erbgut der Rundhaube
verankert ist, kann er nach Kreuzungen in nachfolgenden Generationen
aufgedeckt werden. Bei Lage auf demselben Chromosom und geringer
Distanz sollte es ein sehr seltenes Ereignis sein, hier nach den
Befunden nicht so selten. Die in der F2 und auch nach
Rückpaarungen an die rundhaubige Ausgangsrasse gezogenen Tiere mit
Muschelhaube und Spitzkappe sind meist in der Ausprägung federarmer
als die Ausgangstiere, die Struktur oft im Nacken tiefer angesetzt
und ungeordneter. Auch mähnenartige Erscheinungen treten auf als
Zeichen dafür, dass für die korrekte Ausprägung mehr Faktoren
maßgeblich sind als die hier in den Fokus gestellten potentiell
zwei. Dafür, dass Modifikatoren (Gene an unterschiedlichen Genorten)
verantwortlich sind, und kein Allele (alternative Gene am selben
Genort), sprechen auch die bisherigen molekulargenetische Analysen.
Wie gezeigt fanden Shapiro
u.a. in ihrer Untersuchung zwar Belege dafür, dass das
EphB-Gen (Ephrin receptor B2) bei Reinerbigkeit für alle
unterschiedlichen Haubenformen verantwortlich ist, sie fanden
bislang aber keine Hinweise auf Allele und auch nicht auf andere
Genbereiche, die zwischen Spitz- und Muschelkappe differenzieren
könnten.
Rundhaubige nach der Verpaarung von Spitzkappigen mit Glattköpfigen
Auch den umgekehrten Fall gibt es, dass nach Kreuzungen von
Glattköpfigen mit Spitzkappigen in späteren Generationen Rundhaubige
fallen. Bei Gültigkeit der Modellvoraussetzungen im Fall der
Spitzkappigen aus Kreuzungen von Muschelhaubigen muss die Anlage
latent bei den glattköpfigen Partnern vorhanden gewesen sein, was
bei einigen der in den Büchern ‚Pigeons Genetics‘ und ‚Taubenzucht‘
aufgezeigten Fällen sehr offensichtlich ist.

 
Kreuzung von spitzkappigem Gimpeltäuber und blauer Brieftaube mit
Jungtieren der F1 und, darunter abgebildet, 6 ausgewählte
Jungtiere der F2. Quellen: Sell, Pigeon Genetics und
Sell, Taubenzucht.
In einigen Praxisfällen ist das eine erste und plausible Vermutung.
Mit traditionellen Tests sind diese Vermutungen bei vertretbarem
Aufwand meist nicht aufzuschlüsseln. Es ist eher eine Aufgabe für
zielgerichtete molekulargenetische Analysen.
An anderer Stelle ist ausführlich dokumentiert worden, wie aus einem
spitzkappigen Gimpeltäuber und einer glattköpfigen Hochfliegertaube
bereits in der ersten Generation zur Hälft glatt- und zur Hälfte
spitzkappige Tiere gefallen waren und aus einer umfangreichen F2
eines spitzkappigen Täubers der F1 mit einer
glattköpfigen Täubin der F1 neben Spitzkappigen und
Glattköpfigen einige Jungtiere mit deutlicher Muschelhaube (Sell,
2012).

Spitzkappige und rundhaubiges Jungtier der F2 aus der
Verpaarung eines spitzkappigen Gimpeltäubers mit einer glattköpfigen
Hochfliegertäubin: Quelle: Sell, Pigeon Genetics 2012 und Sell,
Taubenzucht 2019
Paarungen von Spitzkappigen mit Rundhaubigen
Für die genetische Klassifizierung von Phänotypen wichtig die
Beobachtung, dass nicht alle Kreuzungen von Spitzkappigen und
Rundhaubigen, wie in der Untersuchung von
Soederberg,
rundhaubige F1 ergeben. Geschwister können nebeneinander
rundhaubig und spitzkappig sein. Hier aufgezeigt aus der Kreuzung
eines spitzkappigen goldenen Täubers, der aus Goldschwarzflügeln
gefallen ist, mit einer schwarzen muschelhaubigen Täubin.

Muschelhaubige und spitzkappige Nachzucht aus der Verpaarung eines
reinerbigen Spitzkappigen mit einer reinerbigen genetisch nicht
verwandten Muschelhaubigen. Quelle: Sell, Taubenzucht, S. 24.
Über den Tellerrand hinaus
Bei der Analyse komplexer Erscheinungen ist es nützlich, bisherige
Untersuchungen und Testpaarungen zu sondieren und Fakten über
bereits vorgenommene Experimente zur Kenntnis zu nehmen. Dann lassen
sich Übereinstimmungen und Widersprüche zu bekannten
Vererbungsmechanismen feststellen. Nicht in das eigene Denkschema
passende Ergebnisse sollten nicht ausgeblendet werden. Durch eigene
Versuchsanordnungen kann man Wissenslücken füllen, zweifelhaft
erscheinende Ergebnisse erneut prüfen und ggf. eigene Hypothesen
finden, die eine bessere Erklärung der Fakten liefern. Es ist bei
komplexen Merkmalen eine durch Fakten gestützte Aufstellung von
Hypothesen, die durch neue Informationen verworfen, aber auch
verbessert werden können.
Sinnvoll
ist es, diese Hypothesen an unabhängigen Daten erneut zu prüfen und
andere in unabhängige Prüfungen einzubinden. Hypothesenfindung und
Hypothesenprüfung werden dadurch getrennt und die Gefahr verringert,
auf Scheinkorrelationen hereinzufallen.
Fußangeln gibt es bei empirischen Untersuchungen genug. Bei der
ersten Untersuchung durch
Morgan 1911 war es wahrscheinlich eine Fehlklassifikation der
Ausgangstiere (mischerbig und nicht reinerbig) verbunden mit einer
zu geringen Stichprobengröße.
Christie (1877-1927) und
Wriedt (1883-1929) wurden vor den geplanten weiterführenden
Untersuchungen durch den Tod abberufen.
Untersuchungen epistatischer Effekte bei Tauben standen zu der Zeit
auch erst in den Anfängen. Bei Gimpeltauben hat man nicht nur früh
die Vererbung der Kappe studiert, sondern auch den Bronzefaktor.
Erste Tests fanden zwischen Kupfergimpeln und einfarbig schwarzen
und roten Tauben statt. Daraus fielen überwiegend Schwarze, was zur
Annahme eines rezessiven Erbganges des Gimpelbronze führte. Falsch,
wie später erkannt wurde. Schwarze und die meisten Roten haben
genetisch den Farbausbreitungsfaktor ‚Spread‘, und der unterdrückt,
wie rezessives Rot, weitgehend Gimpelbronze. Bei der Paarung mit
Blauen und Blaugehämmerten zeigt sich Bronze in der ersten
Generation, wie beiläufig auf den hier gezeigten Fotos über die
Kreuzung von Spitzkappig und Glattköpfig zu sehen ist. Auch der
Gimpeltaugenphänotyp (gimpel-pattern) mit bronze oder goldenem
Körpergrundgefieder und andersfarbigen Decken und Schwanz tritt in
der ersten Generation in der Regel ein. Bei den frühen Tests wirkte
Spread ‚Verhinderer‘.
Was hat das mit der Kappe zu tun? Es geht nicht um das jeweilige
Merkmal, sondern um die Flexibilität des Denkens. Wenn beim
Gimpelbronze und bei vielen weiteren komplexen Merkmalen
verhindernde oder fördernde Faktoren wirken, dann wäre es
verwunderlich, wenn solche Erbmechanismen bei einem gerade
untersuchten Merkmal ausgeblendet werden könnten. Nicht aufgeklärte
Widersprüche sind immer auch eine Chance, tiefer in die Materie
einzudringen. Und dazu braucht man in vielen Fällen auch ein
Grundverständnis für epistatische Effekte im weitesten Sinn und wird
sich nicht auf einem Grundverständnis der drei Mendelschen Gesetze
ausruhen können.
Literatur:
Aldrovandi, Ylyssis, Ornithologiae, Francofvrti M.DC.X (1610).
Christie, W., und Chr. Wriedt, Die Vererbung von Zeichnungen, Farben
und anderen Charakteren bei Tauben. Zeitschrift für induktive
Abstammungs- und Vererbungslehre 32 (1923), S. 233-298.
Morgan, T.H., Inheritance on crosses between different races of
pigeons, Biological Bulletin Vol. 21 (1911), S. 215-221.
Musavi, Seyyed Mohammad, Book of Pigeons (auf Persisch/Farsi), um
1770.
Sell, Axel, Pigeon Genetics, Applied Genetics in the Domestic
Pigeon, Achim 2012.
Sell, Axel, Taubenzucht. Möglichkeiten und Grenzen züchterischer
Gestaltung, Achim 2019.
Shapiro, M. u.a., Genomic Diversity and Evolution of the Head Crest
in the Rock Pigeon, Science DOI: 10.1126/science.1230422, Published
Online January 31 2013.
Soederberg, Erik, Ein Beitrag zur Genetik der Spitzkappe und der
Rundkappe bei Haustauben, Hereditas Vol. 8 (1927), Issue 3, S.
363-366.
|