|
Schnabellänge bei Haustauben
IV: Ein geschlechtsgebundener Faktor Ku2 bei kurzschnäbligen Haustauben
Beak-Length IV:
A sex-linked trait Ku2 for short beaks
in the Domestic Pigeon
Über die
Bezeichnungen der Schnabellänge bei Haustauben in der Rassetaubenzucht und
über die Grundlagen der Vererbung der Schnabellänge wurde auf dieser
Homepage bereits mehrfach berichtet. Auswertungen der Literatur und
eigene Testpaarungen zeigen, dass kurze Schnäbel der Haustaube durch das
Zusammenspiel von zwei Erbfaktoren erklärt werden können, von denen der
zweite einem geschlechtsgebundenen Erbgang folgt.

Auszug aus dem Buch „Taubenrassen“ 2009.
Geschlechtsgebundene und nicht geschlechtsgebundene Vererbung
In der
Genetik unterscheidet man zwischen Genen, die auf dem Geschlechtschromosom
liegen, und denen, die auf anderen Chromosomen liegen. Bei Tauben werden
z.B. die Grundfarbe mit den Allelen Wild-Typ (schwarz), Dominant Rot und
Braun, die Verdünnungsfaktoren, der Stipperfaktor St, einschließlich der
Allele wie Faded und Qualmond, sowie eine besondere Form der letalen
Schwimmfüßigkeit geschlechtsgebunden vererbt. Die Kurzschnäbligkeit der
Tauben beruht nach jüngsten Ergebnissen wesentlich auf dem Zusammenwirken
von zwei voneinander unabhängigen Genen, von denen das eine ebenfalls auf
dem Geschlechtschromosom liegt. Das andere wird nicht geschlechtsgebunden
vererbt. Eine ungewöhnliche Konstellationen, für die es bei Tauben nach
Kenntnis des Verfassers bisher kein Beispiel gibt. Weitere modifizierende
Faktoren mögen eine Rolle spielen.
Kurzschnäbligkeit in der Literatur
Über die
Vererbung der Schnabellänge wurde auf dieser Homepage schon mehrfach
berichtet. In der Literatur wurde der erste umfangreiche Bericht zur
Vererbung des kurzen Schnabels schon 1923 von den Norwegern Christie und
Wriedt in der Zeitschrift für induktive Abstammungs- und Vererbungslehre
vorgelegt. Die Arbeiten der beiden Autoren wurden von Wexelsen
fortgesetzt, der 1937 im Journal of Experimental Zoology einen Report gab.
Schließlich folgt 1939 ein umfassender Bericht von Harms in der Jenaischen
Zeitschrift für Naturwissenschaft. Die genannten Autoren konnten auf ein
großes Datenmaterial zurückgreifen. Christie und Wriedt hatten z.B. 302
Tiere der Ausgangspopulationen, F1, F2 und
Rückpaarungen ausgemessen. Bei Harms waren es allein bei der Kreuzung von
Schildmövchen mit Dänischen Tümmlern 62 Tiere der ersten Generation und 59
der zweiten. Es wurden bei Rückpaarungen an Schildmövchen 58 Tiere und bei
Rückpaarungen an Dänische Tümmler 53 Jungtiere gezogen.
Trotz der
großen Zahlen waren die Schlussfolgerungen eher vage, was auch daran lag,
dass die Jungtiere nicht nach Geschlecht klassifiziert wurden. Die erste
und zweite Generation zeigten intermediäre Erscheinungen, klare
Spaltungsverhältnisse schienen nicht zu erkennen zu sein. So wurde von
Christie und Wriedt im Wesentlichen ein dominanter Faktor identifiziert,
der die Kurzschnäbligkeit bewirkt, aber angemerkt, dass auch weitere
modifizierende Faktoren eine Rolle spielen könnten. Die Untersuchung von
Wexelsen ist vor allem deshalb interessant, weil er auch relativ
umfangreiches Material über die Kreuzung von Langschnäblern, wie der
Französischen Bagdette, mit Normalschnäblern zur Verfügung hatte und für
Langschnäbler im Verhältnis zu Normalschnäblern zu den gleichen
Schlußfolgerungen kam wie beim Verhältnis der Normalschnäbler zu den
Kurzschnäblern, ein intermediärer Erbgang ohne klare Aufspaltungen.
W.F.
Hollander war der erste, der aus den Daten von Christie und Wriedt
herauslas, dass eine Geschlechtsgebundenheit mitspielen könnte (Origins
and Excursions in Pigeon Genetics 1983). Geschlossen hat er das vermutlich
aus den Angaben, dass aus der Kreuzung von Mövchen-Täubern mit
längerschnäbligen Tümmlertäubinnen im Durchschnitt Jungtiere gefallen
waren, die kürzer als bei der umgekehrten Paarung waren. Larry Chesling
und Gary Fillmore berichteten in den Pigeon Genetics News, Views and
Comments 1986 und 1991 über die Kreuzung kurzschnäbliger Mövchen-Täuber
mit normalschnäbligen Weibchen mit intermediärer Nachzucht. Sie erhielten
allerdings wesentlich kürzerschnäblige Weibchen im Vergleich zu den
Täubern. Sie bekräftigten damit die Vermutung Hollanders, dass
Geschlechtsgebundenheit eine Rolle spielt.
Kurzschnäbler 1,0 x Normalschnäbler 0,1
Die eigenen
Tests waren zunächst nur zur Klärung der Frage gedacht, ob der kurze
Schnabel befriedigend durch den Faktor ku von Christie und Wriedt (ku für
kurz, als dominanter Faktor eigentlich mit Großbuchstaben beginnend „Ku“)
erklärt werden kann oder besser durch das Zusammenspiel zweier oder
mehrerer Faktoren. Wenn man einen nicht geschlechtsgebundenen dominanten
und einen geschlechtsgebundenen partiell dominanten Faktoren unterstellt,
dann werden bei der Paarung eines 1,0 Kurzschnäbler mit einer 0,1
Normalschnäbler Jungtäuber mit der Erbformel Ku1//+; Ku2//+ fallen. Die
Jungweibchen werden geschlechtsgebunden durch die Formel Ku1//+; ●//Ku2
beschrieben. Bei einem partiell dominanten Faktor Ku2 werden die in diesem
Fall hemizygoten Weibchen kürzerschnäblig als die für Ku2 mischerbigen
Täuber sein.
Damit können
auch die Ergebnisse von Paarungen von kurzschnäbligen Mövchentäubern mit
längerschnäbligen Täubinnen, über die Larry Long und Gary Fillmore
berichteten, erklärt werden.
   
Kreuzung von Usbekischem Tümmler mit einer normalschnäbligen einfarbigen
Gimpeltäubin und intermediären Jungtieren. Weibchen (ganz rechts) kürzer
als der Täuber (zweiter von rechts).
Zu denselben
Resultaten führten auch die eigenen Paarungen von kürzerschnäbligen
Tümmlern mit Normalschnäblern.
Normalschnäbliger 1,0 x kurzschnäblige 0,1
Durch
Geschlechtsgebundenheit des 2. Faktors lässt sich auch erklären, warum
beim Verfasser aus einem normalschnäbligen Täuber (24 mm) mit einer
kurzschnäbligen Täubin (16 mm) ausschließlich intermediäre Typen gefallen
sind, die Weibchen allerdings mit einem etwas längeren Schnabel als ihre
Brüder (siehe dazu auch die Abbildungen in früheren Beiträgen zum Thema
auf dieser Homepage). Die Weibchen aus einer solchen Paarung sind bei der
angenommenen Erbstruktur mischerbig für den ersten dominanten Faktor
Ku1//+ und besitzen geschlechtsgebunden den zweiten Faktor nicht. Die
Täuber sind ebenfalls mischerbig für den ersten Faktor, aber auch
gleichzeitig mischerbig für den zweiten Faktor: Ku1//+; +//Ku2. Die
Differenz in der Schnabellänge zwischen Täubern und Tauben ist der Effekt
von Ku2. Dass Ku2 auch in Alleinstellung in Mischerbigkeit einen Effekt
hat, das geht indirekt aus den durchgeführten Rückkreuzungen der Täuber
der ersten Generation an Normalschnäbler hervor, denn andernfalls hätten
daraus wesentlich mehr Jungtiere mit einer gegenüber dem Wild-Typ nicht
verkürzten Schnabellänge fallen müssen.
Hypothese
für die Landkarte des Geschlechtschromosoms
Antwort auf
die Frage der Geschlechtsgebundenheit und nach der Lage des Genes auf dem
Chromosom können Rückpaarungen der F1 und die F2
geben. Geschlechtsgebundene Faktoren sind mit der Grundfarbe gekoppelt,
da diese wie der Verdünnungsfaktor auf dem Geschlechtschromosom verankert
ist.
Der
Ausgangstäuber war ein goldfarbener Kupfergimpel Weißflügel (24 mm). Er
besaß damit auf dem Geschlechtschromosom die Anlage für das Dominant Rot
(BA), den Verdünnungsfaktor Pale und ihm fehlte die Anlage für
Ku2, er entsprach dort dem Wild-Typ +. Das Weibchen wiederum hatte eine
schwarze Grundfarbe (+), war nicht verdünnt (+) und hatte die Anlage für
den kurzen Schnabel Ku2 (16mm). Die Jungtäuber aus diesem Paar haben
diesen Genkomplex geschlechtsgebunden vollständig übernommen. Die
Jungweibchen haben diesen Part nicht übernommen, sondern sind durch das
Chromosom des Vaters geprägt. Über den Lokus der Grundfarbe und den Lokus
der Verdünnungsfaktoren wissen wir, dass diese relativ weit voneinander
entfernt sind, was sich durch die Crossing-Over-Rate belegen lässt
(Hollander 1983). Die Lage auf den Chromosomen könnte die nachfolgend
dargestellte sein.

Wenn kein
Koppelungsbruch auftritt, dann sollten in der F2 alle Jungtiere
mit einer schwarzen Grundfarbe Weibchen sein, den Verdünnungsfaktor nicht
besitzen, und obendrein die Erbanlage Ku2 hemizygot vererbt bekommen
haben (der untere Teil des Chromosomenpaares). Sie haben obendrein die
Chance, reinerbig (25%) oder mischerbig (50%) für den Faktor Ku1 zu sein,
der auf einem anderen und hier nicht gezeigten Chromosom liegt. Das letzte
Viertel besitzt den Faktor Ku1 nicht. Es ist zu erwarten, dass die beiden
Gruppen mit dem Faktor Ku1 so kurz bzw. ähnlich kurz wie die Großmutter im
Schnabel sind.
Die
Ergebnisse der F2 lassen eine ähnliche „Landkarte“ des
Geschlechtschromosoms vermuten, wie sie in der obigen Zeichnung
angenommen wurde. Ku2 scheint enger am Lokus für die Grundfarbe als die
Verdünnungsfaktoren zu liegen. Für die Verdünnungsfaktoren ist bekannt,
dass relativ häufig ein Koppelungsbruch eintritt, woraus geschlossen wird,
dass die Faktoren relativ weit voneinander entfernt liegen.
Die
Koppelung kann durch Koppelungsbrüche aufgehoben werden. Nach dem Bruch
können sich die Teile unterschiedlich zusammensetzen. Interessant sind die
beiden markierten Stellen. Die Möglichkeit mehrfacher Koppelungsbrüchen
wird vernachlässigt, kann aber nicht ausgeschlossen werden. Durch einen
Koppelungsbruch an der ersten (links eingezeichneten) Schnittstelle und
danach folgender Neuformierung des Chromosoms können auch
-
rotfahle
Weibchen mit einem kurzen Schnabel fallen und
-
Pale-Blaue
0,1 mit einem längeren Schnabel.
Durch einen
Koppelungsbruch an der zweiten Schnittstelle und danach folgender
Neuformierung können auch
-
normalschnäblige Rotfahle 0,1 und
-
kurzschnäblige Pale-Blaue/Pale-Schwarze fallen.
Theoretisch
denkbar wäre auch, dass der Lokus für die Kurzschnäbligkeit links vom
Farblokus liegt. Wie man sich selbst klar machen kann, würden in diesem
Fall bei einem Koppelungsbruch keine langschnäbligen Pale-Blauen/Pale-Schwarzen
fallen. Das gilt auch für Rückpaarungen an Normalschnäbler. Wie bei der
Darstellung der Testpaarungen noch ausgeführt wird, sind intermediäre
Pale-Blaue in der F2 gefallen, längerschnäblige Pale-Blaue (23
mm) sind zudem bei Rückpaarungen an Normalschnäbler aufgetreten, so dass
die These einer Lage des Kurzschnabellokuns links vom Farblokus verworfen
wird.
Der
empirische Befund in der Rückkreuzungen und der F2
Hinweise auf
die Geschlechtsgebundenheit geben nicht nur die ersten Kreuzungen von
Kurzschnäbler an Normalschnäbler und umgekehrt, sondern auch die
Rückkreuzungen spalterbiger Täuber der ersten Generation an
Normalschnäbler und die Aufspaltung in der F2. In der
Ausgangspaarung wurde ein normalschnäbliger Goldgimpel-Weißflügel (24mm)
mit einer kurzschnäbligen schwarzgetigerten Tümmlertäubin (16mm) gepaart,
woraus die für die folgenden Versuche genutzten Jungtiere gezogen wurden.
Es sind für schwarzes Pigment und Pale spalterbige Rot- bzw. Aschfahle
(20mm) und eine dominant rote goldfahle (pale) Täubin (21mm). Die Täuber
besitzen Ku2 in Mischerbigkeit, die Täubin besitzt den Faktor
geschlechtsbedingt nicht, sie ist wie die Täuber aber spalterbig für Ku1.
Bei den
Rückpaarungen der F1 an normalschnäblige Täubinnen sind vor allem die
Jungtäubinnen von Interesse. Bei einer Koppelung der Grundfarbe mit dem
Faktor Ku2 sind überwiegend kürzerschnäblige blaue bzw. aus dem Aschfahlen
auch schwarze Täubinnen und längerschnäblige goldfarbene Täubinnen zu
erwarten. Bei diesen Paarungen kann man allerdings von der Färbung der
Jungtiere nicht auf das Geschlecht schließen. Er werden auch blaue und
schwarze sowie rot- und goldfahle Jungtäuber fallen. Da die Jungtiere
nicht alle bis zur Geschlechtsreife behalten wurden, ist ein Vergleich
der beiden Gruppe der Täubinnen mit schwarzer und roter Grundfarbe nicht
möglich. Zu erwarten ist dennoch aufgrund der Koppelung, dass sich die
unterschiedliche Schnabellänge der Weibchen mit schwarzer Grundfarbe und
der mit roter Grundfarbe auf den Gesamtmittelwert der beiden Gruppen
auswirkt. Das hat es auch, denn die 20 schwarzen Jungtiere der Rückpaarung
hatten einen Mittelwert von 20,6 und die 13 brieftaubenroten Jungtiere
einen von 22,15 mm.
Die
Erwartungen wurden auch durch die Ergebnisse der F2 bestätigt.
Abb. 1:
Jungtiere der F2 aus Gimpel-Gold (24mm) x Hochfl. (16mm)
|
mm |
15 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
Summe |
|
F2
aus Gimpel-Gold (24mm) x Hochfl. (16mm) |
2 |
2 |
5 |
1 |
11 |
10 |
1 |
|
|
32 |
|
darunter
rot- oder goldfahl |
|
|
|
|
6 |
9 |
|
|
|
15 |
|
darunter
rezessiv weiß |
|
1 |
1 |
1 |
5 |
|
1 |
|
|
9 |
|
darunter
eindeutig 0,1 (blau bzw. pale) |
2 |
1 |
2 |
|
2 |
1 |
|
|
|
8 |
Von 32
Jungtieren waren 9 weiß und damit nicht nach der Grundfarbe zu beurteilen.
15 waren rot- bzw. goldfahl und 6 waren blau und 2 pale-blau. Die 15 Rot-
bzw. Goldfahlen zeigten alle eine intermediäre Schnabellänge, die beiden
Pale-Blauen (20 und 21 mm)und ein weiteres Jungtier der Schwarzgruppe mit
20 mm waren ebenfalls intermediär, während die übrigen 5 der Schwarzgruppe
kürzerschnäblig waren, drei davon so kurz wie die Ausgangstäubin. Wenn
die Pale-Blauen mit berücksichtigt werden, dann ergibt sich bei den
Jungweibchen mit schwarzer Grundfarbe eine durchschnittliche Schnabellänge
von 18 mm, bei den Jungtäubern und Jungtäubinnen mit roter Grundfarbe von
20,6. Das ist nach einem t-Test statistisch signifikant unterschiedlich
auf dem 99%-Niveau.
 
Jungtiere
der F2
Die
Ergebnisse mit der Konzentration kurzschnäbliger Weibchen mit schwarzer
Grundfarbe und längerschnäbligen Tieren bei den Dominant Roten
entsprechen der These des Zusammenwirkens eines partiell dominanten
geschlechtsgebundenen Faktors mit einem dominanten nicht
geschlechtsgebundenen Faktor für den kurzen Schnabel.
Rückpaarungen der F1 an Normalschnäbler bestätigen das Ergebnis und auch die oben
unterstellte Lage der Gene zueinander auf den Chromosomen. Bei 33
Jungtieren aus der Rückpaarung an Normalschnäbler fielen auch 4 Pale-Blaue/Pale-Schwarze.
Zwei Weibchen davon hatten eine Schnabellänge von 23 mm und entsprachen
damit knapp Normalschnäblern, so dass davon auszugehen ist, dass sie nicht
den Faktor Ku2 besaßen. Das zeigt einen Bruch an der ersten Schnittstelle
im obigen Schaubild an. Für eine genauere Lagebestimmung der Genorte wäre
umfangreicheres Datenmaterial erforderlich.
Schlussfolgerungen für die Praxis
Die
Ergebnisse sind durchaus auch für die praktische Zucht von Interesse. Zum
einen zeigen sie, dass sich das Merkmal Kurzschnäbligkeit nach Kreuzungen
mit Normalschnäblern schnell durch Rückkreuzungen wieder etablieren und
andererseits sich auch schnell aus einem Stamm Normalschnäbler entfernen
lässt. Wenn aus einer normalschnäbligen Rasse positive Eigenschaften,
seien es Vitalität oder andere Eigenschaften, in einen Stamm Kurzschnäbler
eingeführt werden sollen, dann ist es zweckmäßig, ein Weibchen der
Fremdrasse für die Einpaarung zu nutzen. Wenn andererseits aus einer
Kurzschnäblerrasse positive Eigenschaften übertragen werden sollen, dann
sollte man besser ein kurzschnäbliges Weibchen verwenden.
A
sex-linked trait Ku2 for short beaks
in the Domestic Pigeon
Beak length in the domestic pigeon was first analysed by the Norwegian
Christie and Wriedt, Wexelsen and the German Harms in the 20s and 30s of
the last century. Christie and Wriedt proposed the Symbol ku for short
beak (German short = kurz), however, since they did not get the classical
ratios according to Mendel’s laws in the F2 and
their backcrosses they also thought a polygenetic influence possible. Sex
linkage was not considered in the reports by the quoted authors. W.F.
Hollander from the data given by Christie and Wriedt was the first to
presume a sex-linked influence. The guess from Hollander was confirmed by
reports from Larry Chesling and Gary Fillmore. From
shortbeaked Owl-Cocks and Wild-Type hens they got intermediate offspring,
the hens, however, considerable shorter than the cocks (Genetics News,
Views and Comments 1986 and 1991). The result was confirmed by crosses of
a short beaked Tumbler cock with Wild-type hens by the author.
From the data published in literature and additional test matings by the
author the conclusion is derived that a short beak is caused by the
combined effect of a dominant and not sex linked trait Ku1 and a second
one. The second trait Ku2 is sex linked and in heterozygous cocks only has
a minor effect compared to hemizygous hens that have only one member of a
chromosome pair rather than the usual two.
Since colour and dilution factors are sex-linked, an analysis of the
backcross of a F1-cock
from a normal beaked Gimpel cock (dominant red BA, pale dP)
and a short beaked Tumbler hen (Wild-Type +, intense +, Ku2) to Wild-type
and the F2 of
the Gimpel/Tumbler-cross can help to get an understanding of the location
at the sex chromosome. The following mapping of the sex-chromosome is
assumed.
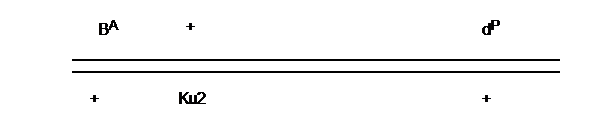
Results for the F2 from
a Gimpel cock (beak length 24 mm) and a Tumbler hen (16mm) are given in
Table 1. Unfortunately recessive white (9 individuals) troubled the
analysis, therefore only 8 hens could be clearly sexed by their colour.
However, the predominance of short beaked individuals in this group and
the predominance of intermediate individuals in the group of dominant reds
(15 cocks and hens) confirm the existence of the proposed linkage.
Figure 1: F2 from light bronze (gold) Gimpel cock (24mm) x
Highflyer hen (16mm)
|
mm |
15 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
total |
|
F2
total |
2 |
2 |
5 |
1 |
11 |
10 |
1 |
|
|
32 |
|
dominant red in both sexes (pale and intense) |
|
|
|
|
6 |
9 |
|
|
|
15 |
|
recessive white in both sexes |
|
1 |
1 |
1 |
5 |
|
1 |
|
|
9 |
|
blue and blue-pale hens |
2 |
1 |
2 |
|
2 |
1 |
|
|
|
8 |
From the backcross of a
F1 cock
to Wild-Type also some pale-blue individuals with a long beak were raised.
If we exclude multiple crossing-over as unlikely their appearance support
the assumed rough mapping of the sex-chromosome and indicate a
crossing-over between the colour locus and the locus for beak length.
|