|
It
is not all dominant or recessive. The discovery and documentation of
inheritance mechanisms in pigeon colours
In 1865 Gregor Mendel
presented his 'Experiments on Plant Hybrids'. From the observation
of characteristic differences in traits and subsequent crosses,
backcrosses and splits, he postulated the 'Mendelian laws' of
heredity. The results did not reach pigeon genetics until after
1900.
Validity of Mendel's laws in pigeons
In 1905 Loisel tried
without success to trace the check pattern back to Mendel's laws.
Probably irritated by the fact that he also had individuals with red
basis colour and grizzles in his stock and with the knowledge of
that time could not classify the genetic basis of the combinations.
In 1911 Bonhote and
Smalley showed the
validity of Mendel's laws also in pigeons in the pattern with the
dominance of checks over bars. In 1914
Cole showed that the
recessive red in tumblers also followed Mendel's rules.
|
 |
Check pattern x bar pattern results in
heterozygous check offspring, which split in the next
generation according to Mendelian rules. |
Epistatic effects
In 1907, Bateson
coined the term epistasis after experiments with the flower colour
of peas. A recessive or dominant gene completely or partially
suppresses the effect of a non-allelic other gene.
In 1914, Cole showed
that black pigeons differed from those with pattern by a hereditary
factor. The dominant spread factor has a covering (epistatic) effect
over pattern. In 1922, Sarah
van Hoosen Jones demonstrates the largely epistatic effect of
spread on extensive data when analysing the pattern. 1928
Metzelaar shows the
epistatic effect of recessive red over the basic colours
|
 |
Homozygous Black (Spread) x Blue Bar
gives black offspring. When mated with each other, the
pattern hidden under black are revealed in some of the
offspring. Here the check pattern. |
Sex-linked inheritance
In 1908, Doncaster
demonstrated the concept of sex-linked inheritance for the first
time in animals using variants of the gooseberry peeper.
In 1911 Bonhote and
Smalley and finally Cole
in 1912 demonstrated
that the mechanism of sex-linked inheritance also applied in pigeons
to the mating of blues and the dilute colour silver (dilute blue).
Thus, the phenomenon that silvers from blue parents were always
females - which had already puzzled
Darwin - had received
a resolution. In 1912 Cole
showed that this hereditary mechanism also applied to the mating of
blacks with dilute dun-coloureds and of reds with dilute yellows.
The difference between dominant and recessive red, was documented
together with Kelly in
1919. Here they also showed the sex-linkage of dominant red.
|
 |
Homozygous dilute cocks (here yellows) x
non-dilute hemizygous females (here reds) result in
hemizygous yellow females and heterozygous red cocks |
The inheritance of Pigeons with the genes reduced,
Carl Graefe 1951, and
Rubella, Gerhard Knopf
2002, follow the same rule.
|
 |
Rubella-cock x non-rubella hen (upper
line) results in cocks heterozygous rubella, and hemizygous
rubella hens. |
Hereditary sexual colour dimorphism in pigeons
1868 Darwin discusses
the possibilities of creating strains with colour differences in
cocks and hens. As an example, he finds wine-red pouters in the
literature, in which only the cocks would show black spots. Not
explainable at that time, according to the analyses of
Hawkins (1931) they
were heterozygous cocks for black and dominant red basic colour.
In 1925, the Norwegians
Christie and Wriedt
analysed the dominant sex-linked stipper gene in Danish Tumblers.
Indirectly they show the characteristic whitish colouring of
homozygous cocks.
|
 |
Homozygous stipper pigeons are whitish in
colour and usually have health deficits in contrast to
hemizygous females (here a pair from the author's breeding). |
1942 Hollander shows
hereditary sexual dimorphism in faded with the possibility of
creating reproducible auto-sex strains (Texan Pioneers)
|
 |
Sexual dimorphism in ash red faded
Texans. The lighter-coloured homozygous cocks reproduce
when mating with hemizygous frosty females |
1970s: TIM KVIDERA discovers in the USA at Racing Homers a gene
'Frosty' probably mutated from Faded. Pure-bred cocks have more
distant resemblance to faded heterozygous cocks and hemizygous faded
hens on a blue color base. Female pigeons corresponded to the wild
type blue-bar. In 1988 he estimated them to be a recessive
sex-linked allele of Stipper and Faded. In 2000 ANDREAS LEIß shows
that Thuringian Selfs have the same hereditary behaviour. The
females, corresponding to the blue-ground-coloured cocks, are
blue-bar. After outcrossing upon other breeds there are occasionally
slight lightenings in front of the tail band. This is also found in
heterozygous faded cocks and is described by BECHSTEIN 1807 in the
historical 'tail pigeon' native to Thuringia as giving its name.
Unlike Reduced, Rubella, the dilution factors, and also Faded, the
hemizygous females in Frosty do not show their sex-linked factor.
|
  |
Thuringian blue-ground-colored cock and a
black non-frosty hen with black and blue bar offspring. At
the right a heterozygous cock and a hemizygous frosty hen.
|
Genetic Linkages
1911 to 1929 Morgan
and his co-workers make extensive Drosophila mappings of chromosomes
and document frequencies of linkage breaks in genes located together
on a chromosome
1919 Cole and
Kelly empirically
determine a cross-over rate (frequency of linkage breaks) for the
sex-linked primary colours and dilution factor of 40% (50% would be
the statistical expectation for independently inheriting genes)
|
 |
Cross-over test by a cock heterozygous
for dominant red and dilute on the same chromosome to a
hemizygous dilute coloured dominant red (thus yellow) hen.
Source: Sell, Taubenzucht 2019 |
1938 Hollander
determines a close linkage with a cross-over rate of 2-3% for the
non-sex-linked factors recessive opal and pattern.
Completion of the basic colours
In 1924 Metzelaar
(quoted from Steele
1931) was the first to recognise brown as an independent basic
colour. That brown is an allele of Dominant Red was shown by
Hawkinson 1931.
|
 |
Brown as sex-linked basic colour.
Homozygous male and hemizygous female do not differ in
colour. |
Additional non-allelic interactions
Following Bateson,
further non-allelic factor interactions are shown after 1907,
especially in plants. Brief introductions easily give the impression
that each trait is caused by one or very few genes. However, almost
all traits are likely to be the result of cumulative or
complementary interaction of several genes (Bartelmess
1965: 724). Pigeon fanciers express this by referring to 'modifying'
factors as an explanation for variations judged to be small. In the
case of larger variations, potentially significant factors are
explicitly named.
1925: That dominant genes override the effect of non-allelic genes
is not surprising for pigeon fanciers. Stipper, for example,
transforms pure recessive reds in English Short Faced Tumblers and
Danish Tumblers into DeRoy. This was shown for Danish Tumblers by
Christie and
Wriedt in 1925.
Fulton (1876) could
not yet classify the colouration in English Short Faced Tumblers.
|
 
|
DeRoy English Short Faced Tumbler (left).
Youngsters DeRoy, Brown Stipper, Kite and red Agate at
Danish Tumblers in the own loft
|
1929 Christie and
Wriedt identify
'bleaching factors'. They only appear on recessive reds that are
bred from heterozygous Brander couples. Recessive red thus is the
prerequisite to show that effect. The bleaching (whitening)
increases with age.
|
   |
Bleaching type I at Christie and Wriedt,
Recessive Reds from Danish Brander. Source: Christie and
Wriedt 1929 |
In 1876, Fulton had
described recessive red pigeon that moult in parts to white as a
complementary colour of the Almond breed. The trait has been
transferred to the Danish Stippers bred from English Short Faced
Tumblers. Here, too, the gene is only found in recessive reds. In
larger experiments Tim
Kvidera (1982) for red Viennese White Shields, and
Andreas Leiß (2008)
for red white shields and Uzbek red Chinny, have confirmed this.
They postulated a white-shield factor, which presupposes homozygous
recessive red.
|
  
|
English Short Faced Tumbler red Agate,
Viennese White Shields recessive red, Chinny Uzbek Flying
Tumbler |
2010, 2011 Hein van Grouw
and Dina Mergeani
found the basic ash red red colour to be a prerequisite for the
naked-neckedness of Romanian Tumblers. This links a feature to a
non-colouring trait.
|
 
|
Romanian Naked-neck Tumbler (photo at
the right: Layne Gardner) |
Heterozygous recessive red has been documented as a modifier to
improve standard colouration for show in breeding almond English
Short Faced Tumblers since
Fulton (1876) and later in Danish Stippers. A positive effect
on the desired standard colouration may explain why the recessive
factor persists in the breed and why recessive red keeps splitting
out.
|
  |
Danish Brown Stipper Cock and Red Agate
from Danish Stippers in nest plumage and shortly before
completion of moult from the author’s loft. ‘Brown’ is a
term of the standard, genetically the basic color is black. |
In Copper and Gold Gimpel pigeons, recessive red has a positive
effect on the desired copper or gold colouring when heterozygous,
analogous to the effect in almonds. From well coloured parents self
reds and golds split out (so already
Goodall 1899).
|
  |
Gold Blackwing Gimpel, heterozygous
recessive red, and Self gold cock (Recessive Red and Pale)
from Gold Gimpel Pigeons from the own loft Source, Sell,
Genetics of Pigeon Colouring (German language), Achim 2015 |
1925 Christie and
Wriedt found that
recessive reds regularly are raised from Danish Brander that moult
white-pied (see above). However, they did not connect this
observation to bronze colouration of Brander. 2008, 2010 and 2011
Ko van Vliet and
others for Dutch Chimney Sweepers,
Wim Halsema and
Bill Peterson for
Danish Brander show that the existence of heterozygous recessive red
leads to the transformation of otherwise kites into 'Brander
bronze'.
|
  |
Danish Brander and Dutch Chimney Sweeper,
heterozygous for Recessive Red |
2012 Interaction of recessive sex-linked genes: Frosty and Rubella.
Both factors are located on the sex chromosome (see above). One
difference is that in Frosty the factor is only shown by homozygous
cocks. In Rubella, the hemizygous females also show it. Based on
Rubella, the Frosty factor, which does not show in heterozygous
cocks and hemizygous hens, has a colour effect. Hemizygous
Frosty-Rubella females turn ice-grey in body plumage. Checks and
bars are weakened in colour. This was also shown by heterozygous
cocks when mating Frosty-Rubella hens with the wild type. Often less
distinct. Pattern are still slightly brownish or only anthracite
toned down. Pure-bred Frosty-Rubella cocks are light silver-grey. In
colour contrast the family corresponds to sex-dimorph coloured
faded. Due to the relatively long distance of the loci for Frosty
and Rubella on the sex chromosome, the connection is fragile. It is
quickly lost in outcrosses.
|
 
|
Homozygous Frosty-Rubella-couple with
distinct sex dimorphism (left), heterozygous frosty-rubella
cock from wild-type x frosty-rubella hen (right) at the
author’s loft. |
Molecular genetic anchoring of genes
In 2004, Charles Lee,
in collaboration with a Canadian research group, published a study
in the context of the analysis of the human genome in which it was
shown that larger sections of DNA were present in all individuals
with different copy numbers (Copy Number Variation CNV). In the
analysis of the human genome, potential connections with diseases
are of particular interest.
2017, 2019, 2020 Domyan,
Shapiro, Bruders et al. present extensive studies on the
anchoring of genes in the genome and investigate differences and
effects, e.g., of CNV for Stipper and alleles of the Stipper-gene.
|
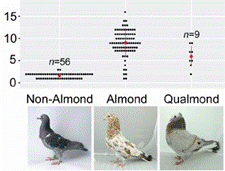 |
Copy number variation at Stipper and
Alleles of the Stippergene. Source: Excerpt from Bruders R,
Van Hollebeke H, Osborne EJ, Kronenberg Z, Maclary E,
Yandell M, et al. (2020) A copy number variant is
associated with a spectrum of pigmentation patterns in the
rock pigeon (Columba livia). PLoS Genet 16(5): e1008274.
https://doi.org/10.1371/journal.pgen.1008274
n=number of tested individuals. |
Literature und Hints on Literature
Hollander, W.F., Origins and Excursions in Pigeon
Genetics, Burrton, Kansas 1983
Levi, W.M., The Pigeon, Sumter South Carolina
1941, revised and reprinted 1969
Sell, Pigeon Genetics. Applied Genetics in the
Domestic Pigeon 528 p., Achim 2012
Sell, Genetik der Taubenfärbungen (German language), 384 p., Achim
2015
Sell, Critical Issues in Pigeon Breeding. What we
know and what we believe to know, Parts I-VI Achim, 60 p., 2020/2021
Introductions in Genetics in English, French
and Dutch
Sell, Introduction to Heredity in Pigeons, 80 p.,
Achim 2022
Sell, Inleiding tot de erfelijkheid bij duiven,
80 p., Achim 2022
Sell, Introduction à l'hérédité chez les pigeons,
80 p., Achim 2022

  

|