|
Pitfalls in traditional and molecular genetic classification of
pigeon breeds
Pigeon breeds can be viewed and subdivided from different
perspectives.
1. historically, striking characteristics and behaviours are
described side by side when depicting domestic pigeons. In
Frisch 1763 and in
other early writings, these got designations for groups of pigeon
breeds such as pouters, tumblers, high-flyers, drummers etc., which
still differed from each other in other details within the group.
These groupings can still be found today as a classification scheme
in the book of standards of the pigeon fancier organisations.
2. the interest of Darwin
(1868) in this context was not directed towards the differences of
breeds and breed groups, but towards the succession of preliminary
stages and preliminary final stages of the development of breeds.
This enabled him to illustrate the rapid change of characteristics
under the influence of selection.
3. even before Mendel
(1865), but increasingly after him, breeds with other
characteristics were seen as a possible source for the improvement
of breeds or for the creation of new breeds by uniting positive
characteristics.
4. molecular genetic classifications of breeds in clusters are made
according to genetic similarities identified by DNA analyses. Breeds
with particularly high similarity to each other, but with a clear
distance to other groups, are grouped together.
Some statements about these studies give the impression that there
is little information about the relationships between breeds,
especially with pigeons:
„as
unlike with many other domestic species, few reliable records exist
about the origins of, and relationships between, each of the breeds”
(Pacheco et al., S.
137).
As explained below, this underestimates the wealth of information
available in the fancier literature. On the other hand, the
possibilities of making unambiguous statements are overestimated in
view of the breeding activities and multiple crossbreeding between
breed groups in the past.
Traditional classifications of breeds according to their utility,
appearance and behaviour
In the historical European literature on pigeons, the description of
characteristics of breeds and their presumed regional origin are
central. So, in Gessner
1555/1557 and in the revised version 1669, in
Aldrovandi around
1600, Willughby 1676,
Moore 1735 and
Frisch 1763.
Frisch had already
distinguished nine basic types and presented one example for each
group pictorially. Field pigeons and Montauben (month pigeons) as
ancestors of today's colour pigeons, drummer pigeons, pouters, owls,
tumblers, Turkish pigeons as representatives of warty pigeons,
Jacobins and fantail pigeons as representatives of the structural
pigeons.
 
Fig. 1: Representatives of two main groups of
domestic pigeons by Frisch
1763, Pouters and Turkish Pigeon (Source:
Sell: Taubenrassen
2009).
In the German book of standards the groups 'form pigeons’ and 'hen
pigeons' have been added. The heterogeneous group 'form pigeons'
also includes the large utility pigeons listed as a group in
Buffon 1772, among
them the giant pigeons already classified as 'Runts' in
Willughby 1676 and as
Romans in Dürigen
1886. In 2022, the breeding stock of the German BDRG shows 35 pouter
breeds from the 'prototype' shown by
Frisch, and far more
than 100 breeds of tumblers and high-flyers. The classification of
the NPA, the American Pigeon Breeders Association, is similar and
follows the tradition of other associations.
Historical classifications may be overtaken by development and prove
inaccurate.
Dixon
had already written this in 1851 for some subgroups of tumblers that
had degenerated from a flying point of view: they would be called
tumblers because they would roll over if they could still fly (p.
118). Associations sometimes react to changes that cannot be
overlooked by regrouping. For example, the Coburg Larks and Lynx
Pigeons, which were classified as coloured pigeons by
Dürigen in 1886, have
become form pigeons in the current book of standards, and Strasser
as former hen pigeons have also become form pigeons.
Darwin: Changes and emergence of breeds by selection
For Darwin, pigeon
breeds were important because he was able to use them as examples to
show the rapid change of characteristics through selection. It is
about the ancestors of these breeds and about the preforms that
still existed at his time. It is not the side by side of breed
groups that is of interest, but in this context the differences
between the breeds and their predecessors (parent stock). These are
indicated in his illustrations in dotted lines. He was able to show
the extreme change of characteristics in a relatively short time. In
the 'family tree' of the English Short Faced Tumbler, for example,
it is shown with a small figure and extremely short beak as the end
of a developmental path. Among its direct ancestors are the Common
Tumbler, common in England, and, more distantly, the related Lotan
Tumbler and the Persian Tumbler.
 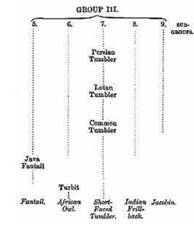
Fig. 2: English Short Faced Tumbler and its line of
development in Darwin
1868 (Source: Sell,
Taubenrassen 2009).
The Carrier, with a long beak and relatively high and slender
stance, is also at the end of an evolutionary line. Darwin had
measured 3.6 cm beak length from the tip to the corner of the mouth
in extreme specimens. The Dragon (now Dragoon), the Bussorah Carrier
and other Carrier breeds with much shorter beaks are mentioned as
predecessors in Darwin’s
illustration (cf. Sell,
2009, p. 124).
 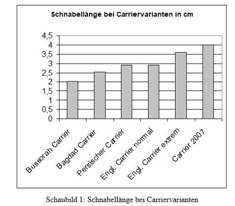
Fig. 3: English Carrier in
Darwin 1868, and
development of beak length over time (Source:
Sell, Taubenrassen
2009).
Whether the development of the beak length in both breeds was due to
selection alone remains to be seen. For in the historical literature
there are indications and evidence that the present French Bagdette,
which existed under other names at that time, was involved in the
development of the English Show Carrier, and, in the case of the
beak length of the English Short Faced Tumbler, also owls.
Aldrovandi
in Europe did not know the fantail in 1600. In 1669 it appears in
the version of Gessners
revised by Horst as
Cprian peacock tail. It was fetched from Holland by local noble
lovers for a lot of money. The number of tail feathers is given as
26 instead of the 12 of a normal domestic pigeon. This is also the
number quoted by Darwin with reference to
Willughby (1676).
Moore (1735) names 38,
Buffon (1772) 32 and
Boitard and
Corbié (1824) 42. In
England, according to Darwin,
it was not so much a question of the number, but also of the general
posture (general carriage). Today, this has changed even further
with extremely short backs (Fig. 4).
Willughby
(1676) describes them with at least 26 tail feathers instead of the
normal 12, Moore
(1735) with 38 and Buffon
(1772) with 32.
Darwin
quotes Boitard and
Corbiè (1824) who
counted 42. In England, however, it was not so much about the
number, but also about the general posture (general carriage). This
has changed even further today with extremely short backs according
to
Darwin
(Fig. 4, 10).
 
Fig. 4: Fantail at
Darwin
and development from Buffon
(1772) via P.J. SELBY
(1835/1843) to today on the cover of the book Taubenrassen.
Creation of new breeds and transmission of traits across breed
groups
The fact that traits - positive and negative - are hereditary was a
firm knowledge of practical animal breeding even before
Mendel. Under the
keyword "Livestock Breeding" in the Book of Inventions, Leipzig and
Berlin 1864, we find the following statements:
"In relation to performance ... the individual species, horse, goat,
cattle, etc., are not only, but also individual branches of these
species among themselves again very different. The name breed means
as much as a variety distinguished by special characteristics common
to all members of the breed. We only find breeds among those animals
which man has taken into his immediate environment, made into
domestic animals and accustomed to his service"... "But it is just
as certain ... that man has it in his power to form new breeds
according to his will"... (in German language, p. 204ff.). And
further on breeding strategies: "The animal breeder can now preserve
the existing breeds or change them in a refining way. A breed is
preserved by mating the best animals within it; it is improved by
inheriting forms or characteristics which it lacks for a desired
purpose or in general, namely by mating animals of different breeds
in which those desired characteristics appear sporadically" (Buch
der Erfindungen 1864, p. 204, quoted at
Sell, Taubenrassen, p.
191).
For many pigeon breeds this is documented in detail by
Boitard and
Corbié already in
1824. In Germany, originally inconspicuous drumming pigeons were
adapted to the taste of the breeders by crossing with colour
pigeons. Thus, the newly developed drumming pigeon breeds were long
referred to in the literature as 'bastard drumming pigeons' (Neumeister
1837 and Neumeister /
Prütz 1876).
 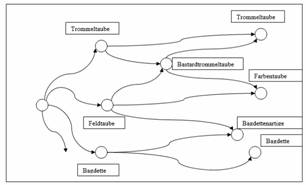
Fig. 5: Drumming pigeons and bastard drumming pigeons in
Neumeister 1837,
Neumeister /Prütz
1876.
Sketch of the interrelationships of breed groups
(Source: Sell,
Taubenrassen 2009).
The group of tumblers and high fliers is particularly receptive to
traits originally associated with other breed groups. High stance,
long necks and long beaks due to influence of French Bagdettes (Fig.
6), short and thick beaks due to Owls, feather ornaments of Drummers
and Fantails, colourings of Colour Pigeons.

Fig. 6: German Magpie Tumblers in
Schachzabel 1910,
Magpie Tumbler in Lyell
1887 and English Magpie Tumbler from 1938, illustrated in
Levi 1969 (Source:
Sell, Pigeon Genetics
2012).
In Magpie Tumblers, pigeons modified by crossbreeding first appeared
as a 'modern type of the breed' before they became accepted as
successors to the old breed. This was also the case with Maltese
Pigeons, Kassel Tumblers and others (Sell,
2009).

Fig. 7: Maltese of old and ‘modern type’ at
Dürigen 1868 and 1906,
source: Sell,
Taubenrassen 2009.
Strasser and Lynx pigeons have also not developed from the pigeons
shown in Lavalle und
Lietze 1905 by
selection in figure and size alone.
 
Fig. 8: Change of the Lynx Pigeon from a Colour
Pigeon to a Form Pigeon and of the Strasser from a Hen Pigeon to a
Form Pigeon (Source: Sell,
Pigeons Genetics. Applied Genetics in the Domestic Pigeon, 2012).
A source of breed mixing is also the transfer of newly discovered
colour factors to other breeds and breed groups. Discovered in 1945
in roller pigeons, for example, the 'Reduced' factor has been
rapidly transferred in the USA to Genuine Homer, Giant Homer, Racing
Homers, Long Faced Tumblers and other breeds. It came to Europe via
such Long-Faced Tumbler crosses. In the Dortmund area the breeding
partnership Christoph Mooren
and Thomas Schmidtmann
created reduced Cologne Tumblers from these. From these the
hereditary factor was transferred by
Norbert Dietrich to
German Modenesers and by
Fritz Muchow to Colour Pigeons in Thuringian Shield Pigeons.
Thomas Voss
transferred the factor to European Racing Homers. In the meantime,
the factor is present in Owls, Drumming Pigeons and other breed
groups.

Fig. 9: Transfer of the Reduced hereditary factor
from roller pigeons in the USA to many breeds within a few years
(Source: Sell,
Critical Issues in Pigeon Breeding, Part IV 2021).
The transfer of other factors can be traced with similar accuracy in
the hobby literature. For example, for the factor indigo uncovered
in crosses in the USA in the 1940s, which was later shown to be
found in high performing European racing pigeon strains as well (Sell
2012, 2019).

Fig. 10: Transfer of the hereditary factor indigo in
a few generations to other breed groups such as high-flyers from
Racing Homers in the author’ loft and Fantails (Source:
Sell Pigeon Genetics.
Applied Genetics in the Domestic Pigeon, 2012).
At the same time as the colour factors are transferred from one
breed to the other, genes from other gene regions are also found in
this and other colour classes through crosses with them.
Molecular genetic classifications
Forms of representation
Following a study by
Stringham et al. (2012), a number of other studies have
appeared on pigeon breed similarity and classification in clusters
and representations in dendrograms. The study used 32 unlinked
microsatellite markers to genetically characterise 361 individuals
from 70 domestic pigeon breeds and two free-living populations.
Similarly extensive, and partly building on it, the study by
Pacheco et al. (2020).
Confidence-building that the tumblers and high-flyers shown here in
the excerpt from the dendrogram given by
Stringham, such as
Birmingham Roller (BIR), Tippler (TIP), West of England Tumbler
(WOE), remain largely together, which confirms or seems to confirm
the assumed commonality of origin.
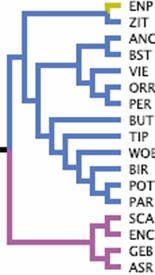
Fig. 11: Example of a dendrogram. Excerpt from
Stringham and others
(2012). A breed classification from the English Cropper (ENP) to the
American Show Racer (ASR).
Similarity by common ancestry or recent crosses?
A proven similarity does not mean a reliable statement about the
origins of the breed. Similarity is determined between current
stock. The similarity may be due to common ancestors, it may also be
due to interbreeding between the breeds studied or between preforms
of these breeds. It is therefore not surprising that in a study
presented by a Polish research group (Biala
et al. 2015), a major finding was that the breeds studied were
subject to permanent genetic mixing.
Lexicographic classification versus classification by measuring
similarity at multiple gene loci.
The newly formed clusters are no substitute for grouping in pigeon
fancy. The English Cropper (ENP) will not be grouped with the
Stargard Shaker (ZIT) in the fancy, despite its grouping in the
dendrogram (Fig. 11). Historically, a single characteristic decides
on the grouping of pouters, drumming pigeons and tumblers/highflyers.
Pouters blow up their crop, drummer have a special vocalisation.
They are still pouters or drummer respectively, even if they differ
in size, colouration, posture and feather ornaments (cap, foot
feathering, etc.). Even with the formerly classified giant pigeons,
only this one criterion decided with the size.
The molecular genetic clustering is different. Here, a
multi-dimensional approach is chosen to define a cluster by
determining the gene regions to be included in the analysis. In
Frisch, genes
responsible for drumming, the inflation of the crop or the tendency
and ability to fly high and tumble were the only ones responsible
for group membership. In molecular genetic clustering, this is in
each case only one trait among many, which is only indirectly taken
into account, if at all.
The importance of the individuals selected for analysis
For the results of the analyses, it is important which individuals
represent the respective races. Dominant hereditary factors, for
example, can be transferred from one breed to another within a few
generations. In the case of recessive ones, it takes somewhat
longer. Even experts of the breed are often unable to detect
differences from the original breed a few generations after
crossbreeding. Individuals of one breed can therefore differ
considerably in the measured genetic distance, which is also pointed
out by Pacheco et al.
in the Supplement for their study. Breeders from Holland presented
some years ago black-winged copper Gimpel Pigeons (Archangel) into
which King Pigeons had been crossed a few generations earlier to
obtain more body mass. Even experts could not find any differences
to the pure breed. Some breeders will consider the first
cross-breeding results of a mating of a peak-crested golden-white
winged cock of the Gimpel Pigeons with a black tigered short-beaked
highflyer hen as differently coloured Gimpel Pigeons. Some of the
plain headed and peak-cested golden white wings originating from the
first back-mating upon the golden-white-wing cock were difficult to
distinguish from pure-bred birds by type, beak length and
colouration, even by experts (Fig. 12).
 
Fig. 12: Mating of a peak-crested golden-white-wing
Gimpel cock with a plain-headed black-tigered highflyer hen with two
of their young. On the right, a female of the first back-mating to
the golden-white-wing cock at the author's loft (source:
Sell, Genetik der
Taubenfärbungen, Achim 2015).
If at first only the genome of birds coming directly from crosses
has changed, these changes will affect the whole breed through
mating with other flocks over the years. Reasons for such
cross-breeding can be colouration, feather ornaments, flight ability
or vitality in general (Sell,
2012, 2019). Especially in rare breeds with frequent breeding
borrowings from other breeds, a molecular genetic finding therefore
only represents a snapshot.
Irritations in the representation of dendrograms
There are various algorithms that lead to different dendrograms and
thus give the viewer different impressions of the network of
relationships between the breeds. Some irritating assignments to
clusters are due to the reasons mentioned above, as
Pacheco et al. also
noted with reference to background information on the animals
studied. Assignments of breeds with large genetic distances to each
other point to still other methodological reasons. So do the cases
where it could be assumed from the small genetic distance that the
breeds would quickly find themselves in a common cluster. When this
does not happen, it may be due to third party relationships. The
environment of the objects also influences the assignment.
Deletions, or even additions of races, can trigger a complete
reorganisation of the dendrogram.
Concluding remarks
For Darwin, the
historical perspective of races and the appearance of preforms
(parent stocks) were of particular importance for showing rapid
changes through selection. He was able to show this exemplarily with
outwardly extremely different breeds.
In pigeon fancying, the differentiation of breed groups was and
still is about a brief characterisation, which was usually based on
a single, easily identifiable characteristic that was considered
significant. As early as
Frisch 1763, these were and still are pouting, drumming,
tumbling, warts, special feather structures, etc. Genetic
similarities in other traits are considered secondary.
One justification for classifying breeds via molecular genetic
studies was that in pigeons, unlike other domestic animal species,
there are few reports on origin and relationships between breeds (Pacheco
et al., p. 137) are not true. The study itself refutes this
by reference to relevant literature and this is shown by the
exemplary references above. Without knowledge of the literature, it
is easy to misinterpret fresh traces from recent interbreeding as
old relationships. Conversely, old ones will be overlooked because
they have been overwritten by more recent crosses. The molecular
genetic analyses obtained are each a snapshot in time. The constant
over time for some breeds is only the breed names. Nevertheless, the
results on genetic differences of breeds remain interesting. In the
medium term, the changes in breeds over time could be monitored. And
this could possibly be done by including historical specimens.
Literature:
Aldrovandi, Ylyssis, Ornithologiæ, Bologna MDC (1600).
Biala, A., et al., Genetic Diversity om eight pure breeds and urban
form of Domestic Pigeon (columba livia var. domestica) based on
seven microsatellite loci Journal of Animal & Plant Sciences, 25(6):
2015, S: 1741-1745
Boitard, Pierre, et Corbié, Les Pigeons de volière et de colombier
ou histoire naturelle et monographie des pigeons domestiques, Paris
1824.
Buch der Erfindungen, Verlagsbuchhandlung Otto Spamer in Verbindung
mit Prof. Dr. Bobrik u.a., Buch der Erfindungen, 3. Band, Leipzig
und Berlin 1864.
Buffon, Georges Louis Leclerc de, Histoire Naturellee, génerale et
particulière, avec las descriptione du cabinet du roi, 1749, Vol. 4
Histoire naturelled des oiseaux, Paris 1772, Pigeons pp. 301ff.
Darwin, Charles, The variation of animals and plants under
domestication. 2 vols. 2nd edn. New York, D. Appleton & Co.
1883. [first published London, John Murray, 1868].
The writings of Charles Darwin on the web
by
John van Wyhe.
Dixon, E.S., The Dovecot and the Aviary, London 1851.
Dürigen, B., Geflügelzucht nach ihrem jetzigen rationalen
Standpunkt, Berlin 1886, Geflügelzucht, 2. ed., Berlin 1906.
Frisch, Johann Leonhard, Vorstellung der Vögel in Deutschlands und
beyläuffig auch einiger fremden, mit ihren Farben…Die Zehnte
Klasse, die Arten der Wilden, Fremden und Zahmen oder Gemeinen
Tauben, Berlin 1763.
Gesner, Conrad, Vogelbuch, Frankfurt am Main 1669, aus dem
Lateinischen mit Verbesserungen durch Georgium Horstium, Reprint
Schlütersche Verlagsanstalt und Druckerei Hannover 1995.
Gesner, Conrad, Vogelbuch. Darin die art/natur und eigenschafft
aller vöglen / sampt jrer waren Contrafactur / angezeigt wirt: ...
Erstlich durch doctor Conradt Geßner in Latein beschriben: neüwlich
aber durch Rudolff Heüßlin mit fleyß in das Teütsch gebracht / und
in ein kurtze ordnung gestelt, Getruckt zu Zürich bey Christoffel
Froschouwer im Jar als man zalt M.D.LVII (1557) (copy).
Lavalle, A., und Lietze, M. (Hrsg.), Die Taubenrassen, Berlin 1905.
Levi, W.M., The Pigeon, Sumter S.C. 1941, revised 1957, reprinted
with numerous changes and additions 1963, reprinted 1969.
Lyell, J.C., Fancy Pigeons, 3rd ed.
London 1887.
Mendel, Gregor Versuche über Pflanzen-Hybriden (1865) von Gregor
Mendel, Vorgelegt in den Sitzungen vom 8.
February und 8. Mach 1865) Net Version.
Moore, J., Pigeon-House. Being an Introduction to Natural History of
Tame Pigeons. Columbarium: or the pigeon house, Printed for J.
Wilford, London 1735.
Neumeister, G., Das Ganze der Taubenzucht, Weimar 1837; Das Ganze
der Taubenzucht., 3. ed. by Gustav Prütz Weimar 1876.
Reprint Verlag Neumann-Neudamm 1988.
Pacheco, G., u.a., Darwin’s Fancy Revised: An Updated Understanding
of the Genomic Constitution of Pigeon Breeds, GBE 2020.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7144551/pdf/evaa027.pdf
Sell, Axel, Taubenzucht. Möglichkeiten und Grenzen züchterischer
Gestaltung, Achim 2019.
Sell, Axel, Pigeon Genetics. Applied Genetics in the Domestic
Pigeon, Achim 2012.
Sell, Axel, Taubenrassen. Entstehung, Herkunft, Verwandtschaften.
Faszination Tauben durch die Jahrhunderte, Achim 2009.
Stringham et al., Divergence, Convergence, and the Ancestry of Feral
Populations in the Domestic Rock Pigeons, Currently Biology (2012),
doi: 10.1016/j.cub.2011.12.045.
Willughby, Francis, Ornithologia, Libres Tres, Londini MDCLXXVI
(1676);
The Ornithology in Three Books. Translated into English, and
enlarged with many Additions throughout the whole work by John Ray,
London 1678.
|