|
Molekulargenetische Studie von Haustaubenrassen
Molecular genetic analysis of the Domestic Pigeon
(see below)
Unter dem Titel „Divergence, Convergence, and the Ancestry of Feral
Populations in the Domestic Rock Pigeon“ geben Wissenschaftler der
Fakultäten für Humangenetik und für Biologie der Universität Utah (Stringham
et al. 2012) einen Bericht über die von ihnen festgestellten genetischen
Verwandtschaften zwischen 70 Haustaubenrassen und zwei freilebenden
Populationen, einer Felsentaubenpopulation in Schottland und einer
verwilderten Haustaubenpopulation in Utah.
Insgesamt wurden 361 Individuen bei 32 nicht gekoppelten Microsatellite
Markierungen untersucht. Das sind eindeutig identifizierbare, kurze
DNA-Abschnitte. Aus den gewonnen Informationen lassen sich bei einem
paarweisen Vergleich Ähnlichkeiten bzw. Distanzen zwischen Populationen
errechnen. Als Indikator für die genetische Distanz wird oft, und auch
hier, der sog. Dest-Wert
ermittelt, der einen Einblick in Unterschiede in Anzahl und Häufigkeit von
Allelen an den betrachteten Genorten in den Populationen vermitteln soll.
Die
Daten können auch aufbereitet werden, um eine Gruppierung der betrachteten
Rassen nach ihrer genetischen Ähnlichkeit vorzunehmen. Auch das ist in der
Studie für eine Untergruppe an Rassen erfolgt. Eine Einteilung in Cluster
kann zunächst sehr grob stattfinden, etwa die Unterteilung in nur zwei
Gruppen, die auf nachfolgenden Stufen weiter untergliedert werden.
Wenn
nur zwei genetisch unterschiedliche Gruppen (K= 2) gebildet werden, dann
verbindet die eine in der Untersuchung äußerlich so unterschiedliche
Rassen wie Kropftauben, Pfautauben und Perückentauben. Die zweite Gruppe
besteht wesentlich aus Tümmlern, dazu kommen Mövchen, Warzentauben u.a.
Diese Gruppe umfasst auch die Brieftauben und die Brieftaubenverwandten.
In Übereinstimmung mit der vermuteten Abstammung der Brieftauben finden
sich Mövchen und verschiedene Brieftaubenrassen auch bei K = 4 und darüber
hinaus in derselben Gruppe. Cumulets zeigen auch bei K = 7 eine
Ähnlichkeit mit Brieftauben und mit Warzentauben (S. 1).
Bei
der gewählten Cluster-Methode werden
·
Mövchen mit dem African Owl (Einfarbiges Mövchen), Orientalische Mövchen,
das Altdeutsche Mövchen und auch – gering abgesetzt – die Chinesentaube,
früh als Gruppe betrachtet, überraschend wird aus
·
europäischen Felsentauben und Modena eine kleine Gruppe gebildet,
·
Feldtaubenartige im weiteren Sinn wie Lockentauben, Archangel (Gimpel)
Englische Trommeltauben, Eistauben, Altholländische Kapuziner, eng
verbunden mit Perückentauben, Schmalkaldener Mohrenköpfe, Sächsische
Mönchtauben und Sächsische Flügeltauben bilden eine größere Gruppe, die
noch einmal etwas tiefer gegliedert ist. Eine weitere Gruppe umfasst
·
Amerikanische Show Racer, die Deutsche Schautaube, Englische Carrier und
Bagdetten. Eine große Gruppe bilden
·
Tümmler, Bodenpurzler, Portugiesische, West of England, Tippler,
Budapester, davon etwas abgesetzt, Persische, Orientalische, Wiener
Mittelschnäblige und, eng miteinander verbunden, Berliner Kurze und
Altstämmer. Am Rande der Tümmlergruppe findet sich der Stargarder
Zitterhals, der überraschend eine größere Gemeinsamkeit mit dem Englischen
Kröpfer zeigt, der wiederum etwas abgesetzt von den übrigen Kröpfern
erscheint.
·
Kröpfer bilden ebenfalls eine Gruppe, wobei Pommernkröpfer und
Norwichkröpfer eine große Gemeinsamkeit zeigen. Horseman, Brünner Kröpfer,
Voorburger Schildkröpfer, Gaditano-Kröper und der Marchenerokröper wurden
ebenfalls in der Gruppe verortet, wobei Voorburger mit Brünnern sowie
Gaditano mit Marchenero jeweils besonders eng zusammenhängen. Abgesetzt
von dieser Gruppe, aber eher dieser, als den anderen Gruppen zugehörig,
finden sich
·
Indische Pfautauben und die Pfautaube.
Die
Distanzen zwischen Rassen sind nicht weniger interessant als der Versuch
der Clusterung und vielleicht sogar aussagekräftiger als die Ergebnisse
bei unterschiedlichen Cluster-Algorithmen.
Für
Brieftauben wird die geringste Distanz bzw. die größte Ähnlichkeit (Dest
= 0,006) mit einer Population von verwilderten Haustauben in Utah, USA,
festgestellt. Diese hat sich offenbar aus verwilderten Brieftauben
gebildet. Angaben zur statistischen Signifikanz der ermittelten Distanzen
werden nicht gegeben, sehr geringe Distanzen wie diese könnten aber auch
dem Zufall geschuldet sein, willkürlich gebildete Untergruppe reiner
Brieftauben könnten sich statistisch auch in geringem Umfang voneinander
unterscheiden. Mit einem Wert von 0,013 tauchten danach Tippler auf und
mit 0,021 Show Racer. Horseman und Rafeno-Kröpfer hatten die
nächstniedrigen Distanzen aufzuweisen mit 0,026 bzw. 0,033, was sich
möglicherweise mit einer nicht lange zurückliegenden Wiedererzüchtung der
im Test genutzten Horseman-Tauben durch Brieftauben und Kröpfer und der
Einkreuzung von Brieftauben in Rafeno-Kröpfer plausibel erklären lässt.
Auch für die historische Herkunft der Cauchois aus Kropftauben finden die
Autoren Indikatoren.
Die
Berechnung der Ähnlichkeiten und die Clusterung ergaben einige
überraschende Ergebnisse. So die auch von den Autoren angesprochene Nähe
der Modena und der Felsentaube in der Clusterung. Vielleicht sollte man
eher den Distanzen zwischen den Gruppen vertrauen, denn der gemessene Dest-Wert
ist mit 0,107 für den Vergleich der Modena mit den meisten anderen Rassen
relativ, aber nicht absolut niedrig. Er wird zudem durch die Distanz zum
King mit 0,087 und zur Indischen Pfautauben mit 0,097 auch unterboten.
Kreuzungen mit Kingtauben haben zur Verbesserung der Masse von Modena in
der Vergangenheit stattgefunden und die Einkreuzung von Pfautauben mag der
Veränderung von Formen gedient haben. Eine Brieftaube, wie man dem Text
zufolge vermuten könnte, war der ursprüngliche Modeneser auch nie, eher
eine Jagetaube mit dem Zweck, fremde Tauben einzufangen (Lyell, 1881, p.
132). Die Veränderung über die Jahrhunderte mag aus der folgenden
Abbildung deutlich werden.
Figure 1: Ausstellungsmodena und Italienischer Modeneser Triganino und
historische Abbildung aus dem Jahr 1876
  
Abb. 216:
Modena
Gazzi khaki nach 2000; Abb.
217: Modeneser
Fliegetaube (Wright, 1879,
Originalquelle nach Lavalle und Lietze, 1905, S. 672/3, wohl Bonizzi, I
Columbi de Modena, Paaolo Tosschi E.E. 1876); Abb. 218: Triganino Gazzi
Schwarz m. bronze Binden nach 2000
Quelle: Axel Sell, Taubenrassen, Achim 2009, S. 204.
Beim
Pommerschen Kröpfer scheint die Nähe zum Norwichkröpfer (0,034) vom Wissen
über die Herkunft aus der Literatur her durchaus plausibel. Auch die Nähe
zum Horseman (0,015) ließe sich gut durch die Vermutung begründen, dass
dieser aus dem gemeinsamen Verwandten Norwichkröpfer und Brieftauben
wieder erzüchtet wurde. Nicht erklärt wird damit die relativ geringe
Distanz der Pommernkröpfer zu den verwilderten Brieftauben in Utah (0,044)
und zu den Brieftauben (0,068). Möglicherweise ist es auch ein
unwahrscheinliches zufälliges Ergebnis aufgrund geringer Stichprobengröße.
Für
die Brieftauben- und Brieftaubenartigen wurden die aus dem
Literaturstudium gewonnenen Ergebnisse weitgehend bestätigt, wie die
folgende dem Buch „Taubenrassen“ des Verfassers entnommene Darstellung
zeigt, in der die in der genetischen Analyse gefundenen nahen Verwandten
der Brieftaube eine prominente Rolle spielen.
Figure 2: Ursprungsrassen der modernen Brieftaube
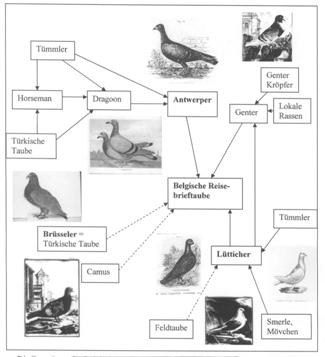
Quelle: Axel Sell, Taubenrassen. Entstehung, Herkunft, Verwandtschaften,
Achim 2009, S. 215.
Die
Autoren zeichnen anhand des ihnen verfügbaren Materials auch die von
Darwin gegebene Unterteilung von 32 Taubenrassen in vier Hauptgruppen und
weitere Untergruppen nach. Sie finden die Aussage von Darwin bestätigt,
dass die äußere Erscheinung zwar generell ein guter Indikator für die
Einschätzung der Verwandtschaft sei, dass in einigen Fällen aber auch
äußerlich sehr unterschiedliche Rassen genetisch eine enge Verwandtschaft
besitzen (Stringham et al., 2012, S. 5). Ein molekulargenetisches
Nachvollziehen der Klassifikation durch Darwin stellt die Clusterung aber
nicht dar. Wie aus Überblicken über die historische Entwicklung von
Taubenrassen deutlich wird, sind bei diesen über Jahrhunderte und bei
einigen Rassen sogar innerhalb einiger Jahrzehnte gravierende Änderungen
durch Kreuzungen eingetreten. Das macht auch die Bildleiste über die
Entwicklung der Modeneser Jage- oder Fliegetaube zu unterschiedlichen
Ausstellungsrassen wie dem Modena deutlich und das zeigen auch die in das
Schaubild über die Entwicklung der modernen Brieftaube eingefassten
historischen Abbildungen. Die Taubenrassen, anhand derer Darwin seine
Klassifizierung vorgenommen hat, existieren in dieser Form nicht mehr.
Molecular
genetic analysis of the Domestic Pigeon
In a report „Divergence, Convergence, and the Ancestry of Feral
Populations in the Domestic Rock Pigeon“ scientists of the University of
Utah (Stringham et al. 2012) present their finding on the relationship
between 70 domestic pigeon breeds and two free-living populations, one of
them wild Rock Pigeons from Scotland and the second ferals in Salt Lake
City, Utah.
The authors used 32 unlinked microsatellite markers (repeating sequences
of 2-6 base pairs of DNA) to genotype 361 individual birds and clustering
software to detect genetically similar individuals in the sample. In the
first step only two genetic clusters were assumed (K=2). The first cluster
combined such morphological different breeds like pouters, fantails and
mane pigeons (Jacobin). The second cluster mainly consisted of the tumbler
breeds (rollers and highflyers), but also owls and wattle breeds. Homers
are included in this group as well.
In accordance with the proposed contribution of carriers, cumulets, and
owl breeds included in this group to the creation of homers, “the owls and
several homer breeds continue to share partial membership in the same
cluster at K = 4 and beyond, and the cumulet shares similarity with the
homers and wattles at K = 7” (ibdi, p. 1f.).
Not all 70 breeds were included in the clustering process and in the
following only a rough impression is given. From the table presented at
p. 6 of the report we may conclude that
·
Owls with African Owl, Oriental Owls, Old German Owls and – slightly
depart – Chinese Owls form a group.
·
European Rock Pigeons and Modena form a second somewhat isolated group.
·
Field pigeon-likes in a wide sense are combined and include Frillbacks,
Archangels, English Trumpeters, Ice-Pigeon, Old Dutch Capuchines near
connected with Mane Pigeons (Jacobin), Schmalkaldener Moorhead, Saxon
Monk, Saxon Wing Pigeon,
·
Homer relatives and wattle pigeon are connected and include the American
Show Racer, German Beauty Homer, English Carrier and Scandaroon.
·
Tumblers form the greatest group and comprise Parlor Rollers, Portuguese,
West of England, Tippler, Budapest Short Faced, and, with some distance
Persian and Oriental Roller, Vienna Medium-face, and – very narrow related
– Berlin Short Faced and Ancient. At the outmost edge of the group the
Stargard Shaker is positioned together with the English Pouter who thus
indicates some distance to the other Pouter breeds.
·
Pouters include Pomeranian, Norwich, Horseman, Brunner, Voorburg, Gaditano
and Marchenero.
·
Fantails and Indian Fantails form a group for its own. The clustering
indicates a more intense relationship to the cropper group than to others.
At least as interesting as clusters are measures of genetic distances
between breeds. And such pairwise Dest-values
and FST-values
were also presented in the annex for breeds with three or more
individuals. For Homers the smallest distance (or the greatest similarity
(Dest
= 0.006) was measured with ferals from Utah. The next lowest distance was
found with Tipplers (0.013), Show Racer (0.021), Horseman (0.026) and
Rafeno-Croppers (0.033). The relationship with Horseman may be explained
by a re-creation of that meanwhile extinct breed by a cross of Homers and
Croppers, and Rafeno-Cropper also may have benefited by different crosses
with homers in the past. The low value for ferals is traced back by the
authors to the influence of wayward racing homers.
There are some surprising results and conclusions. Thus one result of the
clustering-algorithm used was to combine the European Rock pigeon and
Modena in one cluster at the lowest level calculated. Perhaps we should
rely more on the pair-wise distances and the Dest-values.
The value for Modena x Rock-pigeon is lower than for the relation of
Modena with other breeds, but it is not low in absolute terms compared
with other distances, and even the value for Modena x King (0.087) and
Modena x Indian Fantail (0.097) is lower. The relative low value for
Modena x King is not a surprise since King Pigeons are known to have been
uses to improve size, and crosses with Fantails might have been taken
place in the past to shape the figure. The ancient Modena was not a homing
breed, the object of the sport was to let them fly as required, and the
capture of birds of other “Triganieri” (Lyell, 1881, p. 132). The change
of the Modena from the former flying breed is shown above in Figure 1.
The narrow relationship of the Pomeranian Cropper and the Norwich Croppers
(Dest
= 0,034) e.g. seems to be plausible from the information given in old
literature. The low value for the relation Pomeranian x Horseman (0,015)
seems also plausible if we assume that the Horseman was developed from the
common relative of both breeds, the Norwich Cropper. The rather low
distance of Pomeranian Cropper and ferals (0.044) and Homers (0.068) is
puzzling, however.
The conclusions of the study for homers fits well with the theoretical
considerations on the basis of literature as is shown in Figure 2 where
the possible ancestors are included in historical presentations (Sell,
2009).
The authors also try a clustering for the 4 groups of 32 breeds
distinguished by Darwin as far as data were available (14 breeds) and
confirmed the finding that “morphology is a good general predictor of
genetic similarity in domestic pigeons, yet they also show that breeds
that share allelic similarity can be morphologically distinct” (Stringent
et al. 2012, p. 5). In respect to further conclusion we should remember
that all breeds, and also those 12 breeds used in that part of the
analysis are genetically not identical with the breeds and pigeons that
existed in the mid of the 19th century. From literature we know
that several breeds seem to have only the name in common with a breed of
the same name that existed a century before.
Literature:
Darwin, Charles, The variation of animals
and plants under domestication. 2 vols. 2nd edn. New York, D.
Appleton & Co. 1883. [first published London, John Murray, 1868].
Lyell, James C., Fancy Pigeons, London 1881.
Sell, Axel, Breeding and Inheritance in Pigeons, Hengersberg 1994.
Sell, Axel, Taubenrassen. Entstehung, Herkunft, Verwandtschaften.
Faszination Tauben durch die Jahrhunderte, Achim 2009.
Stringham et al., Divergence, Convergence, and the Ancestry of Feral
Populations in the Domestic Rock Pigeons, Currently Biology (2012), doi:
10.1016/j.cub.2011.12.045.
|