|
Es ist nicht
alles dominant oder rezessiv. Entdeckung und Dokumentation von
Vererbungsmechanismen bei Taubenfärbungen
1865 legte
Gregor Mendel seine
‘Versuche über Pflanzen-Hybriden‘ vor. Aus der Beobachtung
charakteristischer Unterschiede von Merkmalen und anschließende
Kreuzungen, Rückkreuzungen und Aufspaltungen postulierte er die
‚Mendelschen Gesetze‘ der Vererbung. Die Taubengenetik erreichten
die Ergebnisse erst nach 1900.
Geltung der
Mendelschen Gesetze bei Tauben
1905 versuchte
Loisel ohne Erfolg,
die gehämmerte Zeichnung als Unterschied zu Binden auf die
Mendelschen Gesetze zurückzuführen. Wahrscheinlich dadurch
irritiert, dass er auch Tiere mit (dominant) roter Grundfarbe und
Schimmel im Bestand hatte und mit den Kenntnissen zu der Zeit die
genetischen Grundlagen der Kombinationen nicht einordnen konnte.
1911 zeigten
Bonhote und
Smalley die Gültigkeit
der Mendelschen Gesetze auch bei Tauben für die Zeichnungen, woran
Loisel noch
gescheitert war. Hämmerung dominiert über die Binden. 1914 zeigte
Cole, dass auch das
Rezessiv Rot bei Tümmlern den Mendelschen Regeln folgt.
|
 |
Gehämmerte Zeichnung x bindige
Zeichnung ergibt mischerbige gehämmerte Nachkommen, die in
der nächsten Generation nach den Mendelschen Regeln
aufspalten |
Epistatische
Effekte
1907 prägte
Bateson nach Versuchen
mit der Blütenfarbe von Erbsen den Begriff der Epistasie. Ein
rezessives oder dominantes Gen unterdrückt ganz oder teilweise die
Wirkung eines nicht-allelen anderen Gens
1914 zeigte
Cole auf, dass sich
schwarze Tauben von denen mit einer Zeichnung durch einen Erbfaktor
unterschieden. Der dominante Spread-Faktor wirkt überdeckend (epistatisch)
über die Zeichnungen. 1922 weist
Sarah van Hoosen Jones
bei der Analyse der Analyse der Zeichnungen den weitgehend
epistatischen Effekt von Spread an umfangreichen Daten nach. 1928
zeigt Metzelaar den
epistatischen Effekt von rezessiv Rot über die Grundfarben
|

|
Reinerbig Schwarz (Spread) x Blaubindig
ergibt schwarze Nachkommen. Untereinander gepaart wird in
einigen Nachkommen die unter Schwarz verborgene Zeichnung
aufgedeckt. Hier die Hämmerung. |
Geschlechtsgebundene Vererbung
1908 wies
Doncaster einen
geschlechtsgebundenen Erbmechanismus bei dunkler und heller
gefärbten Stachelbeer-Spannern nach. Die Versuchsanordnung entsprach
der von Mendel.
Mendel war durch die
Entdeckung nicht ‚widerlegt‘, aber der Geltungsbereich seiner Regeln
wurde auf nicht geschlechtsgebundene Merkmale eingeschränkt.
1911 zeigten
Bonhote und
Smalley und endgültig
COLE 1912 auf, dass die
geschlechtsgebundene Vererbung auch bei Tauben auf die Verpaarung
von Blauen mit der Verdünntfarbe ‚Silber‘ (blaufahl) zutraf. Damit
hatte die Beobachtung, dass ‚Silber‘ aus zwei Blauen immer Weibchen
waren – was schon Darwin
verwundert hatte – eine Auflösung erhalten. 1912 zeigte
Cole, dass der
geschlechtsgebundene Erbmechanismus auch auf die Verpaarung von
Schwarzen mit verdünntfarbenen Dun und von Roten mit
verdünntfarbenen Gelben zutraf. Der Unterschied zwischen dominantem
und rezessivem Rot wurde 1919 zusammen mit
Kelly dokumentiert.
Dabei wurde auch die Geschlechtsgebundenheit von Dominant Rot
aufgezeigt.
|

|
Reinerbig verdünntfarbene Täuber (hier
Gelbe) x Nicht-Verdünntfarben hemizygote Weibchen (hier
Rote) ergeben hemizygote gelbe Weibchen und heterozygote
rote Täuber |
Tauben mit den
Erbfaktoren Reduced, Carl
Graefe 1951, und Rubella,
Gerhard Knopf 2002,
folgen demselben Muster.
|

|
Rubella-Täuber x Nicht-Rubella Weibchen
(oben) ergibt für Rubella spalterbige Täuber und hemizygote
Rubella-Täubinnen. |
Erblicher
Geschlechtsdimorphismus bei Taubenfärbungen
1868:
Darwin diskutiert die Möglichkeiten der Schaffung von Stämmen
mit farblichen Unterschieden zwischen Täubern und Täubinnen. Als
Beispiel findet er in der Literatur weinrote Kropftauben, bei denen
nur die Täuber schwarze Flecken zeigen würden. Damals nicht
erklärbar, nach den Analysen von Hawkins (1931) sind es mischerbige
Täuber für schwarze und dominant rote Grundfarbe.
1925 analysieren die Norweger
Christie und
Wriedt das
geschlechtsgebundene dominante Stippergen (St) bei Dänischen
Tümmlern. Indirekt zeigen sie dabei die Kennfarbigkeit mit weißlich
gefärbten reinerbigen St-Täubern.
|

|
Homozygote Stipper-Täuber sind weißlich
gefärbt und haben in der Regel gesundheitliche Defizite im
Unterschied zu hemizygoten Weibchen (hier ein Paar aus der
Zucht des Verfassers) |
1942
Hollander zeigt den erblichen Geschlechtsdimorphismus bei
Faded mit der Möglichkeit der Schaffung von zuchtfähigen Auto-Sex
Stämmen.
|

|
Geschlechtsdimorphismus bei rotfahlen
faded Texanern. Die helleren Täuber reproduzieren sich bei
der Paarung mit hemizygoten frosty Weibchen |
1970er Jahre:
Tim Kvidera entdeckt
in den USA ein wahrscheinlich aus Faded mutiertes Gen ‚Frosty‘.
Reinerbige Täuber haben entferntere Ähnlichkeit mit mischerbigen
Faded-Täubern und hemizyoten Faded-Täubinnen auf blauer Grundlage.
Täubinnen entsprachen dem Wild-Typ. 1988 schätzt er sie als ein
rezessives geschlechtsgebundenes Allel von Stipper ein. 2000 zeigt
Andreas Leiß, dass
Thüringer Einfarbige dasselbe Erbverhalten haben. Die den
Blaugrundfarbigen entsprechenden Weibchen sind blaubindig. Nach
Fremdkreuzungen gibt es gelegentlich leichte Aufhellungen vor der
Schwanzbinde. Das findet man auch bei mischerbigen Faded-Täubern und
wird von Bechstein
1807 bei der historischen in Thüringen beheimateten ‚Schweiftaube‘
als namensgebend beschrieben. Anders als bei Reduced, Rubella, den
Verdünnungsfaktoren, und auch bei Faded, zeigen die hemizygoten
Täubinnen bei Frosty den Faktor nicht.
|
  |
Thüringer ‚blaugrundfarbiger‘ Täuber
mit schwarzem Weibchen und schwarzen und blauen Jungtieren.
Rechts heterozygoter Jungtäuber und hemizygote Jungtäubin.
Quelle: Sell, Genetik der Taubenfärbungen. |
Genetische
Koppelungen
1911 bis 1929:
Morgan und seine
Mitarbeiter erstellen umfassende Drosophila-Kartierungen von
Chromosomen und dokumentieren Häufigkeiten von Koppelungsbrüchen
bei gemeinsam auf einem Chromosom liegenden Genen
1919
Cole und Kelly
ermitteln empirisch eine Cross-Over-Rate (Häufigkeit von
Koppelungsbrüchen) für die geschlechtsgebundenen Grundfarben und den
Verdünnungsfaktor bei Tauben von 40% (50% ist die statistische
Erwartung bei unabhängig vererbenden Genen).
|

|
Cross-Over Test durch einen für
Dominant Rot und Verdünnung auf demselben Chromosom
mischerbigen Täuber an eine hemizygote verdünntfarbene
dominant rote (damit gelbe) Täubin. Die dunfarbene Täubin
ein Cross-Over. Quelle: Sell, Taubenzucht 2019 |
1938
Hollander ermittelt für die nicht geschlechtsgebundenen
Faktoren Rezessiv Opal und die Zeichnungen eine enge Koppelung mit
einer Cross-Over-Rate von 2-3%.
Vervollständigung
der Grundfarben
1924:
Metzelaar (zitiert nach
Steele 1931) erkannte
als erster, dass Braun eine eigenständige Grundfarbe ist. Dass Braun
ein Allel von Dominant Rot ist, zeigte 1931
Hawkinson.
|

|
Braun als eigenständige Grundfarbe.
Reinerbige Täuber und hemizygote Weibchen entsprechen sich
in der Färbung |
Weitere
nicht-allele Genwirkungen
An
Bateson anschließend werden nach 1907 weitere nicht-allele
Faktorinteraktionen aufgezeigt, vor allem bei Pflanzen. Bei
Kurzdarstellungen entsteht leicht der Eindruck, jedes Merkmal sei
durch ein oder sehr wenige Gene verursacht. Fast alle Merkmale
dürften aber das Ergebnis kumulativen oder komplementären
Zusammenwirkens mehrerer Gene sein (Bartelmess
1965: 724). Taubenzüchter bringen das mit dem Verweis auf
‚modifizierende‘ Faktoren als Erklärung für als gering eingeschätzte
Variationen zum Ausdruck. Bei größeren Abweichungen werden
potentiell bedeutsame Faktoren explizit benannt.
1925: Dass dominante Gene die Wirkung
nicht-alleler Gene überlagern, ist für Taubenzüchter nicht
erstaunlich. Stipper z.B. verwandelt reinerbig Rezessiv Rote bei
Englischen Short Faced Tümmlern und Dänischen Tümmlern in DeRoy. Das
zeigten für Dänische Tümmler
Christie und Wriedt.
Fulton (1876) konnte
die Färbung bei Englischen Short Faced Tümmlern noch nicht
einordnen.
|
 
|
DeRoy Englischer
Short Faced Tümmler (links). Jungtiere DeRoy,
Braunstipper und die Komplementärfarben Kite und Rot Agate
bei Dänischen Tümmlern der eigenen Zucht
|
1929:
Wriedt und Christie
stellen fest, dass Rezessiv Rote, die aus Dänischen Brandern fallen,
mit der Mauser einen ‚Bleaching‘-Faktor zeigen und mit jeder Mauser
stärker weiß ausmausern. Der Bleaching-Faktor wirkt sich ohne
Rezessiv Rot nicht aus.
|
  
|
Verblassung Typ I bei Christie und
Wriedt, Rezessiv Rote aus Dänischen Brandern. Quelle:
Christie und Wriedt 1929 |
Ein zu Weiß Ausmausern
rezessiv roter Tauben hatte
Fulton 1876 für ‚Agates‘ als Nebenfarbe der Almondzucht
beschrieben. Das Merkmal ist auf die aus Englischen Short Faced
Tümmlern erzüchteten Dänischen Stipper übertragen worden. Auch hier
zeigt sich das Gen nur bei Rezessiv Roten. In größeren Versuchen
haben das Tim Kvidera
(1982) für rote Wiener Weißschilder, und
Andreas Leiß (2008)
für rote Weißschilder und Usbekische rote Tschinny, bestätigt. Sie
haben einen Weißschildfaktor postuliert, der Rezessiv Rot in
Reinerbigkeit voraussetzt.
|
  
|
Englischer Short Faced Tümmler rot
Agate, Wiener Weißschilder Rezessiv Rot, Tschinny
Usbekischer Flugtümmler |
2010, 2011:
Hein van Grouw und
Dina Mergeani stellen
die Grundfarbe Dominant Rot als Voraussetzung für die
Nackthalsigkeit der Rumänischen Nackthälse fest. Damit wird ein die
Färbung betreffender Faktor mit einem anderen Merkmal verknüpft.
|
 
|
Rumänischer Nackthalstümmler dominant
rot und gelb (Foto rechts: Layne Gardner) |
Rezessiv Rot in Mischerbigkeit sind als
Modifikator zur Verbesserung der Standardfärbung für Ausstellungen
bei der Zucht almondfarbener Tauben seit
Fulton (1876) bei
Englischen Short Faced Tümmlern und später bei Dänischen Stippern
dokumentiert. Eine positive Wirkung auf die gewünschte
Standardfärbung kann erklären, warum sich der rezessive Faktor in
den Rassen hält und immer wieder Rezessiv Rote ausspalten.
|
  |
Dänischer Tümmler ‚Braunstipper‘ und
Rot-Agate aus Dänischen Stippern im Nestgefieder und kurz
vor Abschluss der Mauser. Quelle, Sell, Genetik der
Taubenfärbungen, Achim 2015 |
Bei Kupfer- und Goldgimpel-Tauben wirkt sich
Rezessiv Rot in Mischerbigkeit analog zur Wirkung bei Almonds
positiv auf die gewünschte Kupfer- bzw. Goldfärbung aus. Aus gut
gefärbten Tieren fallen einfarbige rezessiv Rote und Goldene (so
schon Goodall 1899)
|
 
|
Gold-Schwarzflügel Gimpel und einfarbig
Gold (Rezessiv Rot und Pale) aus Goldgimpeln. Quelle, Sell,
Genetik der Taubenfärbungen, Achim 2015 |
Die Beobachtung, dass aus Dänischen Brandern
regelmäßig Tauben mit dem ‚Bleaching Faktor‘ fielen, hatten schon
Christie und
Wriedt
1929 gemacht, ohne es
mit der Bronzefärbung in Verbindung zu bringen. Dass die
Mischerbigkeit zur Erzielung der Branderbronze-Färbung erforderlich
ist, wurde 2008 und kurz danach für Niederländische Schornsteinfeger
(Ko van Vliet u.a.)
und 2012 für Dänische Brander (Bill
Peterson, Wim Halsema) gezeigt. Die Bedeutung des ‚Bleaching-Faktors‘
bleibt offen.
|
 
|
Dänische Brander und Niederländische
Schornsteinfeger, mischerbig für Rezessives Rot |
2012 Interaktion rezessiver
geschlechtsgebundener Gene: Frosty und Rubella. Beide Faktoren
liegen auf dem Geschlechtschromosom (s.o.). Ein Unterschied besteht
darin, dass bei Frosty der Faktor nur von reinerbigen Täubern
gezeigt wird. Bei Rubella zeigen ihn auch die hemizygoten Täubinnen.
Auf der Grundlage von Rubella hat der gegenüber dem Wild-Typ bei
Täubern und hemizygoten Täubinnen nicht sichtbare Frosty-Faktor
einen Farbeffekt. Hemizygote Frosty-Rubella Täubinnen werden im
Körpergefieder eisgrau. Hämmerung und Binden werden farblich
abgeschwächt. Das zeigten, meist weniger deutlich, auch Täuber bei
der Verpaarung von Frosty-Rubella-Täubinnen mit dem Wild-Typ.
Zeichnungen sind noch leicht bräunlich oder nur anthrazit
abgeschwächt. Reinerbige Frosty-Rubella Täuber sind hell silbergrau.
Im Farbkontrast entspricht die Familie kennfarbigen Faded. Durch die
relativ weite Entfernung der Loci für Frosty und Rubella auf dem
Geschlechtschromosom ist die Verbindung fragil. Sie geht bei
Fremdkreuzungen schnell verloren.
|
 
|
Reinerbiges Paar Frosty-Rubella mit
ausgeprägtem Geschlechtsdimorphismus (links), Jungtäuber
mischerbig Frosty-Rubella aus Wild-Typ x Frosty-Rubella
(rechts ). |
Molekulargenetische Verankerung von Genen
2004 veröffentlicht
Charles Lee in
Zusammenarbeit mit einer kanadischen Forschungsgruppe im Rahmen der
Analyse des menschlichen Genoms eine Studie, in der gezeigt wird,
dass größerer Abschnitte der DNS in allen Individuen mit
unterschiedlicher Zahl an Kopie vorhanden waren (Copy Number
Variation CNV). Bei der Analyse des menschlichen Genoms
interessieren vor allem potentielle Zusammenhänge mit Krankheiten.
2017, 2019, 2020
Domyan, Shapiro, Bruders
u.a. legen umfangreiche Studien zur Verankerung von Genen im Genom
vor und untersuchen u.a. Unterschiede und Auswirkungen von CNV für
Stipper und Stipper-Allele
|
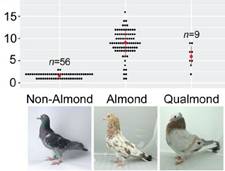
|
Copy number variation bei Stippern und
Allelen des Stippergens. Quelle: Auszug aus Bruders R, Van
Hollebeke H, Osborne EJ, Kronenberg Z, Maclary E, Yandell
M, et al. (2020) A copy number variant is
associated with a spectrum of pigmentation patterns in the
rock pigeon (Columba livia). PLoS Genet 16(5):
e1008274.
https://doi.org/10.1371/journal.pgen.1008274
Anmerkung: n=Zahl der jeweils
getesteten Tauben. |
Literatur und
Literaturhinweise in
Hollander, W.F., Origins and Excursions in Pigeon Genetics, Burrton,
Kansas 1983
Levi, W.M., The Pigeon, Sumter South Carolina
1941, revised and reprinted 1969
Sell, Critical Issues in Pigeon Breeding. What we
know and what we believe to know, Parts I-VI, 60 S., Achim,
2020/2021.
Sell,
Genetik der Taubenfärbungen, 384 S., Achim 2015.
Sell, Pigeon Genetics. Applied Genetics in the
Domestic Pigeon 528 S., Achim 2012.
Sell,
Taubenzucht. Möglichkeiten und Grenzen züchterischer Gestaltung, 368
S., Achim 2019.
Sell,
Verständnis und Missverständnisse in der Taubenzucht. Anekdotische,
unterhaltsame und lehrreiche Anmerkungen zu offenen Fragen, Teile
I-VI, 60 S., Achim 2020/2021.
Einführende Literatur:
Sell, Introduction to Heredity in Pigeons, 80 p.,
Achim 2022
Sell, Inleiding tot de erfelijkheid bij duiven,
80 S., Achim 2022
Sell, Introduction à l'hérédité chez les pigeons,
80 S., Achim 2022
https://taubensell.de

  

|