|
Fallstricke bei traditioneller und molekulargenetischer
Klassifikation von Taubenrassen
Herkunft und Verwandtschaften von Taubenrassen
Taubenrassen können aus verschiedener Perspektive betrachtet und
untergliedert werden.
1. Historisch werden bei der Darstellung von Haustauben auffallende
Merkmale und Verhaltensweisen nebeneinander beschrieben. Bei
Frisch 1763 und in anderen frühen Schriften wurden das
Bezeichnungen für Gruppen von Taubenrassen wie Kropftauben, Tümmler-
und Hochflieger, Trommler etc., die sich innerhalb der Gruppe in
anderen Details noch voneinander unterschieden. Diese Gruppierungen
finden sich bis heute als Gliederungsschema in den
Musterbeschreibungen der Taubenzüchterorganisationen.
2. Das Interesse von
Darwin (1868) war
in diesem Kontext nicht auf das Nebeneinander von Rassen und
Rassegruppen gerichtet, sondern auf das Nacheinander von Vorstadien
und vorläufigen Endstadien der Entwicklung von Rassen. Damit konnte
er die schnelle Veränderung von Merkmalen unter dem Einfluss der
Selektion veranschaulichen.
3. Schon vor Mendel(1865),
aber verstärkt nach ihm, wurden Rassen mit anderen Merkmalen als
mögliche Quelle für die Verbesserung von Rassen oder zur Schaffung
neuer Rassen durch Vereinigung positiver Merkmale angesehen.
4. Molekulargenetische Klassifikationen von Rassen in Clustern
erfolgen nach durch DNS-Analysen festgestellten genetischen
Gemeinsamkeiten. Zusammengefasst werden Rassen mit besonders großer
Ähnlichkeit untereinander, aber mit einem deutlichen Abstand zu
anderen Gruppen. Einige Aussagen zu diesen Studien erwecken den
Eindruck, es gäbe gerade bei Tauben wenig Informationen über die
Beziehungen von Rassen zueinander: „as
unlike with many other domestic species, few reliable records exist
about the origins of, and relationships between, each of the breeds”
(Pacheco et al., S.
137). Daher müsse und könne man die Beziehungen aus den
molekulargenetischen Befunden rekonstruieren. Wie nachfolgend
ausgeführt, wird damit die Fülle der in der Liebhaberliteratur
vorhandenen Informationen unterschätzt. Dagegen werden die
Möglichkeiten zu eindeutigen Aussagen angesichts der züchterischen
Aktivitäten und vielfachen Kreuzungen zwischen den Rassegruppen in
der Vergangenheit überschätzt.
Traditionelle Klassifikationen von Rassen nach ihrem Nutzen, nach
Erscheinung und Verhalten
In der historischen europäischen Literatur über Tauben stehen die
Beschreibung von Rassen und die vermutete regionale Herkunft im
Mittelpunkt. So bei Gessner
1555/1557 und in der überarbeiteten Fassung 1669, bei
Aldrovandi um 1600,
Willughby 1676,
Moore 1735 und
Frisch in Deutschland
1763. Frisch hatte
bereits neun Grundtypen unterschieden und für jede Gruppe
exemplarisch ein Beispiel bildlich vorgestellt. Feldtauben und
Montauben (Monatstauben) als Vorfahren der heutigen Farbentauben,
Trommeltauben, Kröpfer, Mövchen, Tümmler, Türkische Tauben als
Vertreter der Warzentauben, Perücken- und Pfautauben als Vertreter
der Strukturtauben.
 
Abb. 1: Vertreter
zweier Hauptgruppen der Haustauben bei
Frisch 1763 (Quelle:
Sell: Taubenrassen
2009)
In der deutschen Musterbeschreibungen sind die Gruppen
‚Formentauben‘ und ‚Huhntauben‘ dazu gekommen. Die heterogene Gruppe
‚Formentauben‘ umfasst auch die bei
Buffon 1772 als Gruppe
aufgeführten großen Nutztauben, darunter die schon bei
Willughby 1676 als ‚Runts‘
und bei Dürigen 1886
als Römer klassifizierten Riesentauben. Aus den Kröpfern sind 2022
in der Zuchtbestandserfassung des deutschen BDRG aus dem bei
Frisch dargestellten
‚Prototyp‘ inzwischen 35 Rassen geworden, bei den Tümmlern und
Hochfliegern weit über 100. Die Klassifizierung des NPA, des
amerikanischen Taubenzüchter-Verbandes, ist ähnlich und bewegt sich
in der Tradition anderer Verbände.
Historische Einordnungen können sich überleben. Das hatte
Dixon schon 1851 für
einige aus fliegerischer Sicht degenerierte Untergruppen der Tümmler
geschrieben: Sie würden Tümmler genannt, weil sie sich überschlagen
(tümmeln) würden, wenn sie noch fliegen könnten (S. 118). Verbände
reagieren auf nicht übersehbare Veränderungen zum Teil durch
Umgruppierungen. So sind z.B. die bei
Dürigen 1886 noch
als Farbentauben klassifizierten Coburger Lerchen und Luchstauben in
der aktuellen Musterbeschreibung zu Formentauben geworden, Strasser
von Huhntauben auch zu Formentauben.
Darwin: Veränderungen und Entstehung von Rassen durch Selektion
Für Darwin waren
Taubenrassen wichtig, weil er an ihnen exemplarisch die schnelle
Veränderung von Merkmalen durch Selektion aufzeigen konnte. Es geht
um Vorfahren dieser Rassen und um die zu seiner Zeit zum Teil noch
existierenden Vorformen. Nicht das Nebeneinander von Rassegruppen
interessiert, sondern in diesem Kontext die Unterschiede der Rassen
von ihren Vorformen (parent-stock). Diese sind in seinen Abbildungen
in gepunkteten Linien angedeutet. Aufzeigen konnte er die extreme
Veränderung von Merkmalen in relativ kurzer Zeit. Im ‚Stammbaum‘ des
Englischen Short Faced Tümmlers z.B. wird dieser mit kleiner Figur
und extrem kurzem Schnabel als Ende eines Entwicklungsweges
aufgezeigt. Unter seinen direkten Ahnen ist der in England
verbreitete ‚Common Tumbler‘ und, weiter entfernt, der verwandte
Bodenpurzler (Lotan Tumbler) und der Persische Tümmler.
 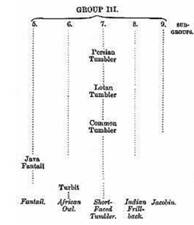
Abb. 2:
Englischer Short Faced Tümmler und seine Entwicklungslinie bei
Darwin 1868 (Quelle:
Sell, Taubenrassen
2009).
Der Carrier, mit einem langen Schnabel und relativ hohem und
schlanken Stand, steht auch am Ende einer Entwicklungslinie. Darwin
hatte in extremen Exemplaren 3,6 cm Schnabellänge von der Spitze bis
zum Mundwinkel gemessen. Als Vorformen in seiner Abbildung genannt
der Dragon (heute Dragoon), der Bussorah-Carrier und andere in
Schnabel wesentlich kürzere Carrier-Rassen (vgl.
Sell, 2009, S. 124).
 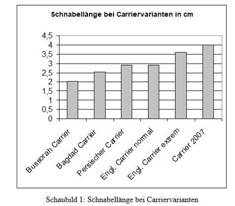
Abb. 3: Englischer Carrier bei
Darwin 1868 und
Entwicklung der Schnabellänge über die Zeit (Quelle:
Sell, Taubenrassen 2009).
Ob die Entwicklung der Schnabellänge bei beiden Rassen allein der
Selektion zu verdanken war, sei dahingestellt. Denn in der
historischen Literatur gibt es Anzeichen und Belege dafür, dass die
damals unter anderen Namen vorhandene heutige Französische Bagdette
bei der Entwicklung des Englischen Ausstellungs-Carrier beteiligt
war, und, bei der Schnabellänge
des Englischen Short Faced
Tümmlers, auch Mövchen.
In Europa kannte Aldrovandi
die Pfautaube 1600 noch nicht. 1669 taucht sie in der durch
Horst überarbeiteten Fassung des
Gessners als Cprischer
Pfauenschwanz auf. Sie sei für viel Geld von hiesigen vornehmen
Liebhabern aus Holland geholt worden. Die Schwanzfederzahl wird mit
26 angegeben, statt der 12 einer normalen Haustaube. Das ist
auch die von Darwin mit Hinweis auf
Willughby (1676)
zitierte Zahl. Moore
(1735) nennt 38, Buffon
(1772) 32 und Boitard
und Corbié (1824) 42.
In England ginge es nach
Darwin aber nicht so sehr um die Zahl, sondern auch um die
allgemeine Körperhaltung (general carriage). Diese hat sich bis
heute mit extrem kurzen Rücken noch weiter verändert (Abb. 4, Fig.
10).
 
Abb. 4: Pfautaube bei Darwin
(links) und Entwicklung von
Buffon (1772) über
P.J. Selby (1835/1843)
bis heute auf dem Cover des Buches Taubenrassen.
Schaffung neuer Rassen und Übertragung von Merkmalen über
Rassegruppen
Dass Eigenschaften – positive und negative – erblich sind, war schon
vor Mendel eine feste
Erkenntnis der praktischen Tierzucht XE "Tierzucht" . Unter dem
Stichwort „Viehzucht“ im Buch der Erfindungen, Leipzig und Berlin
1864, finden wir folgende Ausführungen:
„In Beziehung auf die Leistungen … sind die einzelnen Arten, Pferd,
Ziege, Rindvieh u.s.w. nicht nur, sondern auch einzelne Zweige
dieser Arten unter sich wieder sehr verschieden. Der Name
Rasse bedeutet so viel
als eine durch besondere, allen Gliedern der Rasse bleibenden
Eigenschaften ausgezeichneten Abart. Rassen treffen wir bloß unter
denjenigen Thieren, welche der Mensch in seine nächste Umgebung
genommen, zu Hausthieren gemacht und zu seinen Diensten gewöhnt
hat“… „Es ist aber auch ebenso gewiß, … dass es der Mensch in seiner
Gewalt hat, neue Rassen nach seinem Willen zu bilden“… (so in der
zum Teil veralteten Schreibweise auf S. 204ff.).
Und weiter: „Der Thierzüchter kann nun die vorhandenen Rassen
erhalten oder veredelnd verändern. Erhalten wird eine Rasse durch
Paarung der besten Thiere innerhalb derselben; veredelt wird sie
durch Vererbung XE "Vererbung" von Formen oder
Eigenschaften, welche ihr für einen erstrebten Zweck oder im
Allgemeinen fehlen, und zwar durch Paarung von Thieren verschiedener
Rassen, in denen vereinzelt jene gewünschten Eigenschaften
auftreten“ (zitiert nach
Sell, Taubenrassen, S. 191).
Für viele Taubenrassen ist das durch
Boitard und
Corbié schon 1824
ausführlich dokumentiert. In Deutschland wurden ursprünglich
farblich unscheinbare Trommeltauben durch Kreuzungen mit
Farbentauben dem Geschmack der Züchter angepasst. So wurden die neu
entstandenen Trommeltaubenrassen in der Literatur lange als
‚Bastard-Trommeltauben‘ geführt (Neumeister
1837 und Neumeister/Prütz
1876).
 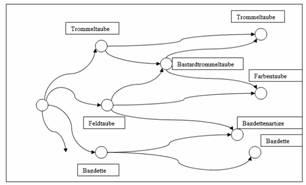
Abb. 5: Trommeltauben und Bastard-Trommeltauben bei
Neumeister
1837, Neumeister/Prütz
1876. Skizze der Zusammenhänge von Rassegruppen (Quelle:
Sell, Taubenrassen
2009)
Die Gruppe der Tümmler und Hochflieger ist besonders aufnahmebereit
für Merkmale, die ursprünglich mit anderen Rassegruppen verbunden
waren. Hoher Stand, lange Hälse und lange Schnäbel durch Einfluss
Französischer Bagdetten (Abb. 6), kurze und dicke Schnäbel durch
Mövchen, Strukturen von Trommeltauben und Pfautauben, Färbungen von
Farbentauben.

Abb. 6: Deutsche Elstertümmler bei
Schachtzabel 1910,
Elstertümmler bei Lyell
1887 und Englischer Elstertümmler aus dem Jahr 1938, abgebildet bei
Levi 1969
(Quelle: Sell, Pigeon
Genetics 2012)
Bei den Elstertümmlern erschienen die durch Kreuzungen veränderten
Tauben zunächst als ‚neue Zuchtrichtung‘, bevor sie sich als
Nachfolger der alten Rasse durchgesetzt hatten. So auch bei
Maltesern (Abb. 7), Kasseler Tümmlern u.a. (Sell,
2009).

Abb. 7: Malteser alter und neuer Zuchtrichtung bei Dürigen 1868 und
1906, Quelle: Sell,
Taubenrassen 2009
Auch Strasser und Luchstauben haben sich nicht allein durch
Selektion in Figur und Größe von den bei
Lavalle und Lietze
1905 gezeigten Tauben fortentwickelt.
 
Abb. 8: Veränderung der Luchstaube von einer Farbentaube zu einer
Formentaube und der Strasser von der Huhntaube zur Formentaube
(Quelle: Sell, Pigeons
Genetics. Applied Genetics in the Domestic Pigeon, 2012)
Eine Quelle der Durchmischung von Rassen ist auch die Übertragung
neu aufgedeckter Farbfaktoren auf andere Rassen und Rassegruppen.
1945 bei Rollertauben (Tümmlern) entdeckt, ist z.B. der Faktor ‚Reduced‘
in den USA schnell in Genuine Homer, Giant Homer, Brieftauben, Long
Faced Tümmler und andere Rassen übertragen worden. Nach Europa kam
er über solche Long-Faced Tümmler-Kreuzungen. Im Dortmunder Raum
wurden daraus durch die Zuchtgemeinschaft
Christoph Mooren und
Thomas Schmidtmann
Kölner Tümmler graugesäumt. Von diesen sprang der Erbfaktor bei
Norbert Dietrich auf
Deutsche Modeneser über und bei
Fritz Muchow auf
Farbentauben bei Thüringer Schildtauben (Abb. 9).
Thomas Voss übertrug
den Faktor auf europäische Brieftauben. Vorhanden ist der Faktor
inzwischen bei Mövchen, Trommeltauben und anderen Rassegruppen.

Abb. 9: Übertragung des Erbfaktors Reduced in wenigen Jahren aus
Rollertauben in den USA in viele Rassen (Quelle:
Sell, Critical Issues
in Pigeon Breeding, Part IV 2021).
Ähnlich genau lässt sich auch die Übertragung anderer Faktoren in
der Hobby-Literatur nachverfolgen. So für den in den 1940 Jahren in
den USA bei Kreuzungen aufgedeckten Faktor Indigo, der, wie sich
später zeigte, auch in leistungsstarken europäischen
Brieftaubenstämmen zu finden ist (Sell
2012, 2019).

Abb. 10: Übertragung des Erbfaktors Indigo in wenigen Generationen
auf andere Rassegruppen wie Hochflieger beim Verfasser (aus
Brieftauben) und Pfautauben (Quelle: Sell, Pigeon Genetics. Applied
Genetics in the Domestic Pigeon, 2012)
Mit den Farbfaktoren werden gleichzeitig Gene anderer Genbereiche
übertragen. die sich durch Kreuzungen mit anderen Farbenschlägen
auch in diesen wiederfinden.
Molekulargenetische Klassifikationen
Darstellungsformen
Im Anschluss an eine Studie von
Stringham u.a. (2012)
sind eine Reihe weiterer Studien zur Ähnlichkeit von Taubenrassen
und zur Klassifikation in Clustern und Darstellungen in
Dendrogrammen erschienen. Die Studie verwendeten 32 nicht verknüpfte
Mikrosatellitenmarker, um 361 Individuen aus 70 Haustaubenrassen und
zwei freilebende Populationen genetisch zu charakterisieren. Ähnlich
umfangreich, und teilweise darauf aufbauend, die Studie von
Pacheco u.a. (2020).
Vertrauensbildend, dass die hier im Ausschnitt aus dem bei
Stringham
gegebenen Dendrogramm gezeigten Tümmler und Hochflieger, wie
Birmingham Roller (BIR), Tippler (TIP), West of England Tümmler (WOE),
weitgehend zusammenbleiben, was die unterstellte Gemeinsamkeit der
Herkunft bestätigt oder zu bestätigen scheint.
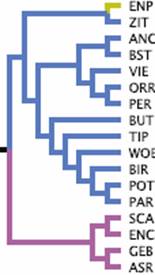
Abb. 11: Beispiel für ein Dendrogramm. Ausschnitt aus
Stringham und anderen
(2012). Eine Rassenzuordnung vom Englischen Kröpfer (ENP) bis zum
Amerikanischen Show Racer (ASR)
Ähnlichkeit durch gemeinsame Abstammung oder kürzliche Kreuzungen?
Eine nachgewiesene Ähnlichkeit bedeutet noch keine verlässliche
Aussage über die Ursprünge der Rasse. Ermittelt wird die Ähnlichkeit
zwischen heutigen Beständen. Die Ähnlichkeit kann an gemeinsamen
Ahnen liegen, sie kann auch an zwischenzeitlichen Kreuzungen
zwischen den untersuchten Rassen oder zwischen Vorformen dieser
Rassen liegen. Es ist daher auch nicht überraschend, dass bei einer
durch eine polnische Forschungsgruppe vorgestellten Studie (Biala
u.a. 2015) ein wesentliches Ergebnis war, dass die untersuchten
Rassen einer permanenten genetischen Vermischung unterworfen waren.
Lexikographische Klassifizierung versus Klassifizierung durch
Messung der Ähnlichkeit an mehreren Genorten
Die neu gebildeten Cluster sind kein Ersatz für die Gruppenbildung
in der Taubenliebhaberei. Den Englischen Kröpfer (ENP) wird man im
Ausstellungswesen trotz seiner Zusammenfassung im Dendrogramm mit
dem Stargarder Zitterhals (ZIT) nicht in eine Gruppe stecken (Abb.
11). Historisch entscheidet bei Kröpfern, Trommeltauben, und
Tümmlern ein einziges Merkmal über die Gruppenzugehörigkeit. Kröpfer
blasen den Kropf auf, Trommeltauben haben eine besondere
Lautäußerung. Es sind auch dann noch Kröpfer bzw. Trommeltauben,
wenn sie sich in der Größe, den Färbungen, der Körperhaltung und in
Federstrukturen (Kappe, Fußbefiederung, etc.) voneinander
unterscheiden. Auch bei den früher klassifizierten Riesentauben
entschied mit der Größe nur dieses eine Kriterium.
Anders bei der molekulargenetischen Clusterung. Bei dieser wird mit
der Festlegung der in die Analyse eingehenden Genbereiche ein
mehrdimensionaler Ansatz zur Festlegung eines Clusters gewählt. Für
das Trommeln, das Aufblasen des Kropfes oder die Neigung und
Fähigkeit zum Hochflug und Tümmeln verantwortliche Gene sorgten bei
Frisch allein für die
Gruppenzugehörigkeit. Bei der molekulargenetischen Clusterung ist
das jeweils nur ein Merkmal unter vielen, das nur indirekt
berücksichtigt wird, wenn überhaupt.
Die Bedeutung der für die Analyse ausgewählten Individuen
Für die
Ergebnisse der Analysen ist wichtig, welche Individuen die
jeweiligen Rassen vertreten. Dominante Erbfaktoren lassen sich z.B.
innerhalb weniger Generationen von einer Rasse auf eine andere
übertragen. Bei rezessiven dauert es etwas länger. Auch Experten der
Rasse ist es oft nicht möglich, wenige Generationen nach Kreuzungen
Unterschiede zur Ausgangsrasse festzustellen.
Individuen einer Rasse können sich in der
gemessenen genetischen Distanz daher wesentlich unterscheiden, was
auch Pacheco et al. im
Supplement zu ihrer Studie aufzeigen. Züchter aus Holland stellten
vor einigen Jahren Schwarzflügel-Kupfergimpel (Archangel) vor, in
die zur Erlangung von mehr Körpermasse wenige Generationen zuvor
Kingtauben eingekreuzt wurden. Auch Experten konnten keine
Unterschiede zur reinen Rasse feststellen. Einige Züchter werden die
ersten Kreuzungsergebnisse (F1) der Verpaarung eines
spitzkappigen Goldweißflügeltäubers der Gimpeltauben mit einer
schwarz-getigerten kurzschnäbligen Tümmlertäubin vom Typ für anders
gefärbte Gimpeltauben halten. Einige der aus der Rückpaarung eines
Weibchens an den Gold-Weißflügel stammenden glattköpfigen und
spitzkappigen Gold-Weißflügel waren vom Typ, Schnabellänge und
Färbung auch von Experten nur schwer von reinerbigen Tieren zu
unterscheiden (Abb. 12).
 
Abb. 12: Paarung eines spitzkappigen Goldweißflügels mit einer
glattköpfigem schwarz-getigerten Tümmlertäubin mit zwei Jungtieren
und rechts davon ein Weibchen der ersten Rückpaarung an den
Goldweissflügel beim Verfasser (Quelle:
Sell, Genetik der
Taubenfärbung, Achim 2015)
Hat sich zunächst nur das Genom bei direkt aus Fremdkreuzungen
stammenden Tieren verändert, so werden sich diese Änderungen durch
Paarungen mit anderen Beständen über die Jahre auf die ganze Rasse
auswirken. Gründe für solche Einkreuzungen können Färbungen,
Federstrukturen, Flugvermögen oder allgemein auch Vitalität sein (Sell
2012, 2019). Insbesondere bei seltenen Rassen mit häufigen
Zuchtanleihen bei anderen Rassen stellt ein molekulargenetischer
Befund daher nur eine Momentaufnahme dar.
Irritationen in der Darstellung von Dendrogrammen
Es gibt verschiedene Algorithmen, die zu unterschiedlichen
Dendrogrammen führen und damit beim Betrachter unterschiedliche
Eindrücke über das Beziehungsgeflecht zwischen den Rassen erwecken.
Einige irritierende Zuordnungen zu Clustern liegen in den vorstehend
genannten Gründe, wie auch
Pacheco et al. mit Hinweis auf Hintergrundinformationen zu
den untersuchten Tieren festgestellt haben. Zuordnungen von Rassen
mit großen genetischen Distanzen zueinander deuten auf noch andere
methodische Gründe hin. So wie auch die Fälle, in denen von der
geringen genetischen Distanz vermutet werden konnte, die Rassen
würden sich schnell in einem gemeinsamen Cluster wiederfinden. Wenn
sie nicht zusammenkommen, kann es an Drittbeziehungen liegen. Auch
die Umgebung der Objekte beeinflusst die Zuordnung. Löschen, oder
auch das Hinzufügen von Rassen, kann eine vollständige
Reorganisation des Dendrogrammes auslösen.
Schlussbemerkungen
Für Darwin waren die
historische Perspektive von Rassen und die Erscheinung von Vorformen
(parent stocks) zum Aufzeigen schneller Veränderungen durch
Selektion von besonderer Bedeutung. Das konnte er exemplarisch an
äußerlich extrem unterschiedlichen Rassen zeigen.
In der Taubenliebhaberei ging und geht es bei der Unterscheidung von
Rassegruppen um eine Kurzcharakterisierung, die meist an einem
einzigen, einfach zu identifizierenden und für bedeutsam gehaltenen
Merkmal festgemacht wurde. Das sind und waren schon bei
Frisch 1763
Kropfaufblasen, Trommeln, Tümmeln, Bewarzung, besondere
Federstrukturen, etc. Genetische Ähnlichkeiten bei anderen Merkmalen
werden als nachrangig betrachtet.
Eine Begründung für die Klassifizierung von Rassen über
molekulargenetische Untersuchungen war, dass es bei Tauben im
Unterschied zu anderen Haustierarten wenig Berichte über Herkunft
und Beziehungen zwischen den Rassen gibt (Pacheco
u.a., S. 137), stimmen nicht. Das widerlegt die Studie selbst durch
Referenz auf einschlägige Literatur und das zeigen die obigen
exemplarischen Hinweise. Ohne Kenntnis der Literatur kann man leicht
frische Spuren durch kürzliche Kreuzungen als alte Beziehungen
fehlinterpretieren. Umgekehrt wird man alte übersehen, weil sie
durch neuere Kreuzungen überschrieben wurden. Die ermittelten
molekulargenetischen Analysen sind jeweils eine Momentaufnahme. Die
Konstante über die Zeit ist bei einigen Rassen nur der Rassename.
Interessant bleiben die Ergebnisse über die genetischen Unterschiede
zwischen Rassen dennoch. Auf mittlere Sicht könnte ein Monitoring
der Veränderungen von Rassen über die Zeit erfolgen. Und das
möglicherweise unter Einbeziehung historischer Präparate.
Literatur:
Aldrovandi, Ylyssis, Ornithologiæ, Bologna MDC (1600).
Biala, A., et al., Genetic Diversity om eight pure breeds and urban
form of Domestic Pigeon (columba livia var. domestica) based on
seven microsatellite loci Journal of Animal & Plant Sciences, 25(6):
2015, S: 1741-1745.
Boitard, Pierre, et Corbié, Les Pigeons de volière et de colombier
ou histoire naturelle et monographie des pigeons domestiques, Paris
1824.
Buch der Erfindungen, Verlagsbuchhandlung von Otto Spamer in
Verbindung mit Prof. Dr. Bobrik u.a., Buch der Erfindungen, 3.
Band, Leipzig und Berlin 1864.
Buffon, Georges Louis Leclerc de, Histoire Naturellee, génerale et
particulière, avec las descriptione du cabinet du roi, 1749, Vol. 4
Histoire naturelled des oiseaux, Paris 1772, Pigeons pp. 301ff.
Darwin, Charles, The variation of
animals and plants under domestication. 2 vols. 2nd edn. New
York, D. Appleton & Co. 1883. [first published London, John
Murray, 1868].
The writings of Charles Darwin on the web
by
John van Wyhe.
Dixon, E.S., The Dovecot and the Aviary, London 1851.
Dürigen, B., Geflügelzucht nach ihrem jetzigen rationalen
Standpunkt, Berlin 1886, Geflügelzucht, 2. ed., Berlin 1906.
Frisch, Johann Leonhard, Vorstellung der Vögel in Deutschlands und
beyläuffig auch einiger fremden, mit ihren Farben…Die Zehnte
Klasse, die Arten der Wilden, Fremden und Zahmen oder Gemeinen
Tauben, Berlin 1763.
Gesner, Conrad, Vogelbuch, Frankfurt am Main 1669, aus dem
Lateinischen mit Verbesserungen durch Georgium Horstium, Nachdruck
durch die Schlütersche Verlagsanstalt und Druckerei Hannover 1995.
Gesner, Conrad, Vogelbuch. Darin die art/natur und eigenschafft
aller vöglen / sampt jrer waren Contrafactur / angezeigt wirt: ...
Erstlich durch doctor Conradt Geßner in Latein beschriben: neüwlich
aber durch Rudolff Heüßlin mit fleyß in das Teütsch gebracht / und
in ein kurtze ordnung gestelt, Getruckt zu Zürich bey Christoffel
Froschouwer im Jar als man zalt M.D.LVII (1557) (Kopie).
Lavalle, A., und Lietze, M. (Hrsg.), Die Taubenrassen, Berlin 1905.
Levi, W.M., The Pigeon, Sumter S.C. 1941, revised 1957, reprinted
with numerous changes and additions 1963, reprinted 1969.
Lyell, J.C., Fancy Pigeons, 3rd ed.
London 1887.
Mendel, Gregor Versuche über Pflanzen-Hybriden (1865) von Gregor
Mendel, Vorgelegt in den Sitzungen vom 8.
Februar und 8. März 1865) Net Version.
Moore, J., Pigeon-House. Being an Introduction to Natural History of
Tame Pigeons. Columbarium: or the pigeon house, Printed for J.
Wilford, London 1735.
Neumeister, G., Das Ganze der Taubenzucht, Weimar 1837; Das Ganze
der Taubenzucht., 3.
Aufl. von Gustav Prütz Weimar 1876. Reprint Verlag Neumann-Neudamm
1988.
Pacheco, G., u.a., Darwin’s Fancy Revised: An Updated Understanding
of the Genomic Constitution of Pigeon Breeds, GBE 2020.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7144551/pdf/evaa027.pdf
Sell, Axel, Taubenzucht. Möglichkeiten und Grenzen züchterischer
Gestaltung, Achim 2019.
Sell, Axel, Pigeon Genetics. Applied Genetics in the Domestic
Pigeon, Achim 2012.
Sell, Axel, Taubenrassen. Entstehung, Herkunft, Verwandtschaften.
Faszination Tauben durch die Jahrhunderte, Achim 2009.
Stringham et al., Divergence, Convergence, and the Ancestry of Feral
Populations in the Domestic Rock Pigeons, Currently Biology (2012),
doi: 10.1016/j.cub.2011.12.045.
Willughby, Francis, Ornithologia, Libres Tres, Londini MDCLXXVI
(1676);
The Ornithology in Three Books. Translated into English, and
enlarged with many Additions throughout the whole work by John Ray,
London 1678.
|