|
Isabell in pigeons and breeding success with guided mate choice
Isabella in old literature
Isabell coloured pigeons are already mentioned in old literature as
a speciality. Neumeister mentions them in pouter pigeons in 1837.
Coloured drawings promoting the colour scheme are first found in
pigeon books in England. Thus, in Tegetmeier 1868 a muffed pair. The
coloured drawing in Fulton 1876 is clearly recognisable as a pouter
pigeon. The delicate colour of the Isabella is emphasised by the
adjacent red and yellow in comparison.


Fig. 1: Isabella pigeons at Tegetmeier 1868 and Fulton 1876
The muffed Isabella Pigeons are named differently in old literature,
sometimes as Dutch, sometimes as Saxon Pouters.
Prütz 1885 shows a clean-legged Brunner Pouter. Behind it in the
drawing a black with white bars. Isabella are often shown in
drawings together with other white-barred ones, as in Fulton's work.
However, the colourations and white bars are genetically based on
different hereditary factors. The fact that they were often shown
together in the picture did not make it easier for the breeders and
later also not for the decoding of the genetic structure.
 
Fig. 2: Isabell-coloured Brunner Pouter and isabell-coloured Prague
Tumbler at Prütz 1885. In the background a light blue with white
bars.
In Prütz's work, next to the Prague isabell-coloured Tumbler, a
light blue with white bars, which is not quite realistic.
Genetically, like the Isabella, it has the hereditary factor
Dominant Opal (Sell 2012, 149-155). The tail bar of the light blue
in reality is bleached to a lighter whitish colour, different from
the drawing.
Breeding problems with Isabella
Despite all the praise, there have always been only a few breeders
of Isabella. This may also be due to ignorance of how to breed this
colouring best. Until the 1960s, however, the breeders did not
mention any breeding problems with Isabella and also not of light
blues with white bars, checks or lacing, which are linked to them by
the common hereditary factor Dominant Opal. Then there were reports
from breeders about poor breeding results. They suspected inbreeding
or diseases as the cause. In the genetics group around W.F.
Hollander in the USA there were notes in the 'Pigeon Genetics
Newsletter' published by him around that time that it was difficult
to obtain purebred Dominant Opals. And if you had an animal that
could be homozygous, there would be vitality problems.
Andreas Leiß from Vienna succeeded in breeding such purebreds in
feral pigeons. He documented them in photos and also showed the
problems of the surviving pure-bred Dominant Opal. Whitish/grey in
colour, not attractive and vitality problems. The reared female died
after a short time, the juvenile cock lived a little longer but did
not reproduce. This matched other reports of grey/white juveniles
already dying in the nest.
Investigation into the reproduction of isabella
It is the merit of Wolfgang Schreiber and his breeding friends of
the Brno and Saxon Croppers to have investigated the thesis of the
failure of the pure dominant opal on a broad data basis. The results
confirmed the earlier assumptions on the lethal or semi-lethal
effect of homozygosity for Dominant Opal (Schreiber 2004). It was
found that from pairs of two isabell-coloured pigeons (648 eggs) on
average 3.2 young were born, from mixed pairs (665 eggs) 5.2. This
could be broken down into a lower number of eggs laid per pair and
lower fertilisation rates in pairs formed by two isabella, lower
hatching rates from the fertilised eggs and higher mortality rates
after hatching before banding (see also Sell 2012).
From Punnett's squares one can read that half of Isabell x Yellow
are expected to be isabell-coloured. That is 2.6 isabella per pair
from 5.2 kittens. From Isabell x Isabell with 3.2 kittens there are
arithmetically just under 2.2 isabella per pair. In percentage terms
it is more, but in absolute terms it is less than in the comparison
pairing.


Fig. 3: Brunner Pouter yellow (complementary color to isabell.
Brunner Pouter isabell (Günter Dietze) and Saxon Pouter isabell
(Dieter Geisemeyer).
Source: Sell, Pigeon Genetics, 2012, photos: Layne Gardner
Instead of yellows, one can also mate with reds as a complementary
colour, if possibly the colouration is also somewhat darker. Spotted
bills are sometimes mentioned as a disadvantage. Epistatically
masked, the reds and yellows should genetically possess the bar
pattern.
Change in the proportion of isabella in the stock without selection
in the case of random mate choice
Lethal genes are quickly lost in nature. This would also happen to a
strain of isabella or light blue pigeons left to itself. If the
breeding stock for the next breeding period is completely replaced
by the offspring, this can be made plausible in fast motion by
simple model considerations. Let's start with a flock of pairs
Isabell x non-Isabell (100% heterozygous Dominant Opal). Directly
readable in Punnett's square, a quarter of the kittens do not have
the factor. Half of the kittens are heterozygous Dominant Opal like
their parents. A quarter of the kittens are homozygous Dominant
Opal. These die already in the egg or shortly after hatching. With
random selection from the young of the year for further breeding,
the percentage of Dominant Opal will have dropped from 100% of the
stock to 66.6%.
When these offspring mate with each other, three combinations are
conceivable. Non-carriers of Dominant Opal with each other (1/9 of
matings), mixed carriers of the gene with each other (4/9 of matings)
and a carrier of the gene with a non-carrier (4/9). Failure by
purebred Dominant Opal will occur in these matings when two trait
carriers mate with each other. If the reproductive behaviour is not
influenced by other confounding factors, the proportion of
reproductive Dominant Opals will be reduced to 50% in this cohort.
If one follows the model's calculation instruction over the
generations, the proportion thins out further to 40%, 33.3%, 29%,
etc. (cf. Fig. 4).
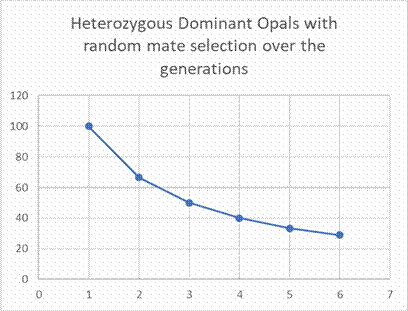
Fig. 4: Model observation of the percentage of heterozygous Dominant
Opals with random mate selection over the generations. Exchange of
parents by offspring after each breeding year.
Stable ratio of 50% of heterozygous individuals in the wild
White-throated Sparrow due to sexual preference for differently
coloured mates (negative disassorting mating)
It is interesting to look for parallels with other animal species.
Frank Mosca recently pointed to studies in the American
White-throated Sparrow (Zonotrichia albicollis). Here, too, there is
a dominant inheritance of the colour variant 'white' over 'tan', the
other variant. By 'white' is meant that the throat patch and a line
above the eye and over the head appear white and not brownish (tan)
(cf. Fig. 5). Here, too, the gene that is considered dominant seems
to reduce vitality in the case of pure heredity, according to the
observations. Why do they still exist, and stable in the population
by 50%?

Fig. 5: White-throated Sparrow, the 'White' variety with white
instead of brown (tan) markings. Source: https://de.wikipedia.org/wiki/Wei%C3%9Fkehlammer#/media/Datei:Zonotrichia_albicollis_CT1.jpg
According to observations on mate choice, mating of white x tan and
vice versa occurred almost exclusively in free-living populations.
As recommended for Dominant Opal in pigeons: Isabell x Yellow and
vice versa. The other matings occurred in the 1 percent range. These
few pairings probably triggered by the absence of other partners. As
with species crosses between domestic and wild pigeons or doves,
there provoked by the keeper. Pure-bred 'whites' were therefore
hardly found in nature, 3 out of 1,989 birds, one male and two
females. The male remained smaller than normal (Hedrick, Tuttle,
Gonser 2018).
In feral domestic pigeons, Johnston and Johnson (1989) had reported
preferences for certain and also different from the own colourations
in mate choice (negative assortative mating), but nowhere near as
clearly as in the White-throated Sparrow. The molecular genetic
difference between Tan and White was found to be chromosomal
inversions involving about 1000 genes on the white-chromosome, with
genes affecting not only colouration but also other characteristics
such as behaviour (ibid.). Thus, whites and tans are differently
aggressive, they also differ in brood care. Among the genes in this
area, presumably also those that directly or indirectly influence
mate choice.
Reactions to lethal genes in pigeon breeding
According to the breeding recommendations for pigeons, in the case
of genes with lethal or semi-lethal effects, two trait carriers
should not be mated with each other, also from the point of view of
animal welfare. The reasons can be seen from the above. In nature,
in some cases, negative assortative mate choice ensures that the
heterogeneity of the initial stock is maintained over the
generations. In pigeon breeding this is simulated by guided matings.
The question of whether there is a natural affinity for divergent
coloured mates in dominant opal-coloured pigeons - similar to the
White-Throated Sparrow - does not seem to have been asked yet.
Whether preferences are signalled by colouration or by other
associated traits remains to be seen. Whether it is better to speak
of recessive or dominant can also be left open for Dominant Opal.
Historically, dominance is spoken of on the basis of colouration. If
the mortality of the young is put in the foreground, the effect is
recessive.
Literature:
Hedrick, Ph. W., Elaina M. Tuttle, and Rusty A. Gonser,
Negative-Assortative Mating in the White-Throated Sparrow.
Journal of Heredity, 2018, 223–231.
Hollander, W.F. (Hrsg.), Pigeon Genetics Newsletter, Volume 1962.
Johnston RF, Johnson SG. 1989. Nonrandom mating in feral pigeons.
Condor.
91:23–29.
Schreiber, Wolfgang, Erhebung über die Nachzucht isabeller Brünner.
Brünner aktuell 2004, 49.
Sell, Axel, Pigeon Genetics. Applied Genetics in the Domestic
Pigeon, Achim 2012.
|