|
Vor 100 Jahren: Sewell Wrights
Inzuchtkoeffizient
Die negativen Wirkungen von Inzucht hatte schon Darwin im 19.
Jahrhundert beschrieben. Ihm, und Tierzüchtern war aber bewusst,
dass eine Einheitlichkeit der Nachkommen und eine Verankerung
positiver Merkmale in einer Rasse ohne Verwandtschaftszucht nicht
möglich ist. Bei kleineren Populationen wie in der Taubenzucht kommt
es automatisch zur Verpaarung engerer Verwandter, zur Inzucht. Die
Frage potentiell negativer Wirkungen bei Reinerbigkeit hatte Sewell
Wright in den 1920er Jahren beschäftigt. Er suchte einen Ausgleich
zwischen den positiven Wirkungen von Reinerbigkeit (Einheitlichkeit
der Nachkommen und Vererbungskraft positiver Merkmale) und den
negativen (Vitalität und Fruchtbarkeit).
Vereinheitlichung der Nachzucht bei konstituierenden Merkmalen wie
Größe, Federstrukturen, Verhalten:
Bei vielen Merkmalen ist Reinerbigkeit erwünscht und aus Erfahrung
für die Lebensfunktionen nicht negativ. Wer glattköpfige Tauben
züchtet, der freut sich nicht, wenn durch Mischerbigkeit für die
Haube gelegentlich ein Jungtier mit Federhaube im Nest liegt. Wo
eine mittlere Körpergröße angestrebt wird, stören in der Nachzucht
Über- und Untergrößen. Reinerbigkeit für die im Standard
festgeschriebene Ausprägung eines Merkmals ist das Ziel.
Fremdpaarungen sind aus dieser Sicht verbunden mit der Gefahr des
Verlustes einer mühsam erreichten Reinerbigkeit des Stammes.
Aufdecken von negativen Merkmalen im Stamm durch
Verwandtschaftszucht:
Durch Inzucht werden auch rezessive vitalitätsmindernde und im
Extremfall letale Faktoren in einem Stamm aufgedeckt.
Schwimmfüßigkeit bis hin zur letalen Schwimmfüßigkeit, Vielzehigkeit
und andere sind aus dieser Sicht das negative Gesicht der Inzucht.
Es ist aber nicht nur negativ, dass sie sich zeigen. Denn
konsequente Züchter haben so die Möglichkeit, über Stammbäume die
genetischen Merkmalsträger zu ermitteln und durch Selektion die
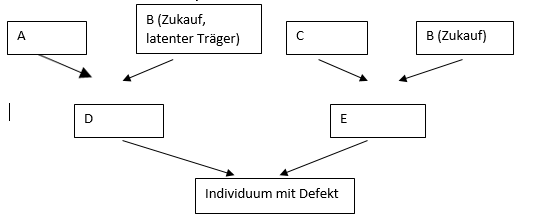
Erscheinung im Bestand frühzeitig zu eliminieren. D und E sind im
Beispiel durch die Abstammung von B latente Träger. Die Geschwister
von D und E mit hoher Wahrscheinlichkeit ebenfalls, wie auch die
Geschwister des betroffenen Individuums.
Äußerlich nicht wahrnehmbare Merkmale
Mit Fortschritten der Molekulargenetik kann man tiefer in das Genom
blicken als mit dem bloßen Auge. So gibt es z.B. unterschiedliche
Allele beim Feder-Keratin Gen und auch beim LDH-Gen, das eine Rolle
bei der Umwandlung von Milchsäure (Laktat) spielt (vgl. den
Überblicksartikel von Eberhard Haase 2021). Zeigte sich hier bei
Untersuchungen bei Brieftauben, dass auch Spitzenflieger nicht
unbedingt reinerbig für dieselben Allele sein müssen, so ergaben
frühere Untersuchungen zu Transferrin-Genotypen, die beim Transport
des in der Leber produzierten Eisens eine Rolle spielen, dass
Reinerbigkeit für bestimmte Allele die Lebensfunktionen negativ
beeinflussen kann. Bei permanenter Inzucht können sich negative
Konstellationen auch bei anderen bisher nicht untersuchten Genen in
Beständen anhäufen und damit das auslösen, was Inzuchtdepression
genannt wird.
Die Berechnung des Inzuchtkoeffizienten
Der von Wright entwickelte Inzuchtkoeffizient berechnet aus einem
Stammbaum, wie im Beispiel dargestellt, den potentiellen Inzuchtgrad
eines Tieres. Potentiell, weil es die Wahrscheinlichkeit bestimmt,
dass zwei Gene eines Genortes abstammungsgleich sind.
Abstammungsgleich bedeutet, dass sie vom selben Ahnen stammen und
über die Generationen weitergegeben wurden.
In dem Beispiel ist es der Ahne B, der sowohl mütterlicher- als auch
väterlicherseits über die Eltern D und E gemeinsam im Stammbaum
steht. Wenn A, B, C keine gemeinsamen Ahnen haben, die
berücksichtigt werden müssten, gilt nach Wright der Koeffizient Fj
für das Individuum j
Fj = Σ (1/2)n1+n2+1, n1 ist dabei die Anzahl
der Generationen vom Elternteil D zum gemeinsamen Ahnen B (hier = 1)
und n2 die Anzahl der Generationen vom Elternteile E zum gemeinsamen
Ahnen B (hier auch = 1). Das Summenzeichen Σ bezieht sich darauf,
dass es mehrere gemeinsame Ahnen geben kann. Dann wird ein Wert für
jeden Ahnen berechnet und anschließend addiert. Für die gezeigte
Halbgeschwisterpaarung errechnet sich ein Inzuchtkoeffizient von
Fj = (1/2)3 = 0,125.
Bei der Verpaarung von Vollgeschwistern wird sich der Wert
verdoppeln, 0,250 (für eine tiefere Darstellung Sell 2012, 2019).
Die Parallelen zu Vererbungsabläufen
Die Formel für den Koeffizienten mag auf den ersten Blick wie Magie
aussehen. Sie hat aber als Hintergrund das gleiche genetische
Verständnis, das auch in den Mendelschen Gesetzen und in der
Darstellung der Punnettschen Quadrate zum Ausdruck kommt.
Unterstellen wir, dass Vollgeschwister miteinander verpaart werden,
deren Eltern genetisch nicht miteinander verwandt sind. Auf einem
Lokus im Genom befinden sich bei einem Elternteil die Allele 1 und 2
und beim anderen die Allele 3 und 4. Bei einer großen Anzahl von
Nachkommen könnten daraus vier Genkombinationen fallen, (1//3),
(1//4) und (2//3) sowie (2//4). Bei einer sehr großen Zahl
zufälliger Vollgeschwisterpaarungen sind in der nachfolgenden
Generation 1/16 der Jungtiere reinerbig für Allel 1. Es sind in der
statistischen Erwartung auch jeweils 1/16 der Jungtiere reinerbig
für die anderen drei Allele, damit insgesamt 4/16 oder ¼ bzw. die in
der Formel von Wright gegebenen 0,250. Für die obige Diskussion der
Inzuchtdepression fast noch bedeutsamer: Die Wahrscheinlichkeit,
dass ein Jungtier der F2 ein Gen, z.B. das Gen 1 nicht
hat, beträgt ein Viertel. Wenn zufällig für die Weiterzucht zwei
solcher Jungtiere miteinander verpaart werden, dann ist in der
Inzuchtline das Allel 1 verschwunden. Die Wahrscheinlichkeit eines
solchen Zusammentreffens beträgt statistisch ¼ x ¼ = 1/16. Das
erscheint gering, gilt aber auch für den endgültigen Verlust eines
der anderen Allele und für andere Gene des gesamten Genoms. Es kommt
über die Generationen zu einer Zunahme an Reinerbigkeit und, als
andere Seite der Medaille, zu einem Verlust an Allelen in der
Familie. Mit den Transferrin-Genotypen hat man Merkmale gefunden,
bei denen das schädlich sein kann. Das wird nur die Spitze des
Eisbergs sein.
Zum Nutzen der Inzuchtkoeffizienten
Die Berechnungen sind nicht vergleichbar mit der
molekulargenetischen Analyse von Individuen, Abstammungen und
Ähnlichkeiten innerhalb von Populationen. Diese waren zur Zeit
Wrights nicht möglich und sind es für Taubenzüchter auch heute kaum.
Bleibt der Koeffizient als ein leichter zu ermittelnder
Anhaltspunkt. Die absoluten Zahlen sind für sich betrachtet nicht so
wichtig, die Unterschiede bei unterschiedlichen Zuchtstrategien
vielleicht umso mehr. Immerhin zeigen sie dem Anfänger, dass es
besser ist, mit zwei Paaren eine Zucht zu beginnen als mit einem
Paar. Der gemessene Inzuchtkoeffizient bei der Verpaarung von
Geschwistern der Nachzucht aus nicht verwandten Eltern untereinander
wird als 0,250 berechnet, bei einer erneuten Geschwisterpaarung
steigt er auf 0,375. Sind Nachkommen eines anderen nicht verwandten
Paares für die Querverpaarung vorhanden, sinkt der gemessene
Koeffizient der Nachzucht der ersten Generation auf null, bei
Querverpaarung steigt er in der zweiten auf moderatere 0,125 und
eröffnet für einige Zeit die Möglichkeit einer weniger engen
Verwandtschaftszucht. (AS)
Literatur:
Haase, Eberhard, Molekulargenetik und Brieftaubenzucht, Die
Brieftaube Nr. 25 vom 26.
Juni 2021, S. 6-10.
Sell, Critical Issues in Pigeon Breeding. What we know and what we
believe to know, Part VI, Achim 2021
Sell, Pigeon Genetics. Applied Genetics in the Domestic Pigeon,
Achim 2012
Sell, Taubenzucht. Möglichkeiten und Grenzen züchterischer
Gestaltung, Achim 2019.
Wright, Sewall, Coefficients of Inbreeding and Relationships, The
American Naturalist Vol. 56, No. 645 (Jul. - Aug., 1922), pp.
330-338.

Eine im Hinblick auf die Rassemerkmale rein gezüchtete Familie von
Thüringer Weißlätzen
 
Defekte wie Schwimmfüßigkeit in einer Familie, die von Zeit zu Zeit
bei Verwandtschaftszucht aufgedeckt wird. Die Schwimmfüße führen
auch zur Verkrümmung der Zehenstellung
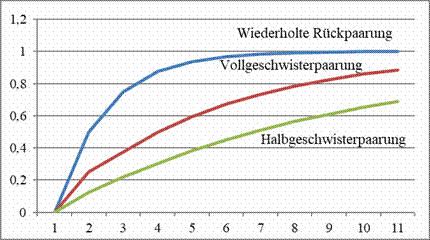
Inzuchtkoeffizienten nach Wright bei unterschiedlichen Paarungen.
Quelle: Sell, Taubenzucht. Möglichkeiten und Grenzen züchterischer
Gestaltung. Zucht und Vererbung in Theorie und Praxis, Achim 2019

Unterschiedliche Inzuchtkoeffizienten. Im Hintergrund durch
Fremdbefruchtung null, im Vordergrund wesentlich höher. Quelle:
Sell, Taubenzucht, Achim 2019
|